 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
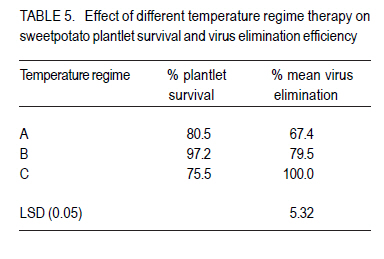
African Crop Science Journal, Vol. 18, No. 3, 2010, pp. 223 - 233 DETECTION AND ELIMINATION OF SWEETPOTATO VIRUSES R. J. RUKARWA, A.B. MASHINGAIDZE1, S. KYAMANYWA and S.B. MUKASA Department of Crop Science, Makerere University, P. O. Box
7062, Kampala, Uganda
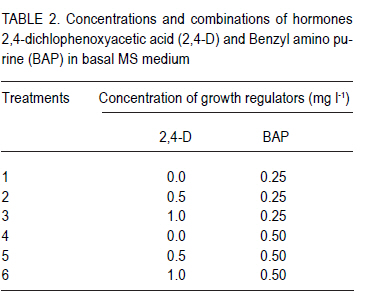
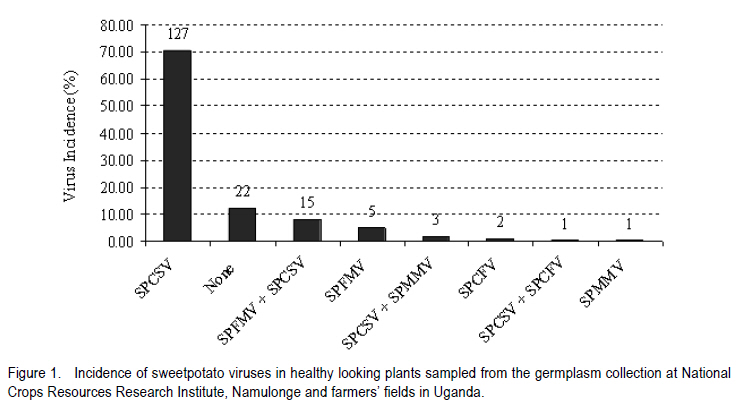
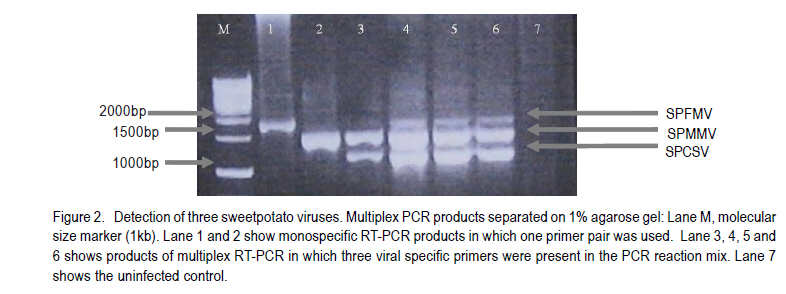
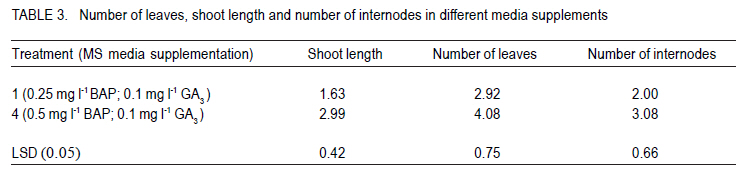
Corresponding author: sbmukasa@agric.mak.ac.ug Code Number: cs10026 ABSTRACT In sub-Saharan Africa, sweetpotato (Impomoea batatas L.) production is greatly constrained by sweetpotato virus disease (SPVD) complex. This study was conducted to assess the incidence of viruses in healthy-looking sweetpotato in Uganda and to optimise modern technologies for virus diagnosis. A collection of healthy-looking sweetpotato vines from central Uganda were serologically assayed for sweetpotato viruses and the positive samples were confirmed by RT-PCR. A multiplex RT-PCR assay was optimised for simultaneous detection of Sweet potato chlorotic stunt virus (SPCSV), Sweet potato feathery mottle virus (SPFMV) and Sweet potato mild mottle virus (SPMMV). The use of in vitro thermotherapy was also investigated as a means of eliminating sweetpotato viruses. Four viruses namely SPCSV, SPFMV, SPMMV and SPCFV were detected mostly as single infections in the healthy looking plants. SPCSV (70. 6%) recorded highest incidence followed by co-infection of SPFMV and SPCSV (8.3%). Based on shoot survival and effectiveness of virus elimination, the best results were obtained by exposing plantlets to daily temperature regime of 32 oC for 8 hr of darkness and 36 oC for 16 hr of light for four weeks. Meristem-tip culture combined with thermotherapy allowed elimination of SPFMV and SPMMV in 77% of plants that were previously infected with the respective viruses. However, elimination of SPCSV was unsuccessful. Key Words: Multiplex RT-PCR, thermotherapy, tissue culture RÉSUMÉ En Afrique sub saharienne, la production de la patate douce (Impomoea batatas L.) est grandement affectée par un complexe de maladies de virus (SPVD). Cette étude était conduite pour évaluer l'incidence maladie des virus sur des boutures apparemment saines de la patate douce en Ouganda et optimiser les technologies pour diagnostic de virus. Des boutures apparemment saines de patate douce collectées au centre de l'Ouganda étaient sérologiquement testées et les échantillons infectés étaient confirmés par RT-PCR. Un essai multiplexe RT-PCR était optimisé pour la detection simultanée du virus du rabougrissement chlorotique de la patate douce (SPCSV), le virus de la marbrure duveteuse de la patate (SPSMV) et le virus de marbrure modérée de la patate douce (SPMMV). L'usage de la thermothérapie in vitro était aussi testé comme moyen d'élimination des virus de la patate. Quatre virus dont SPCSV, SPFMV, SPMMV et SPCFV étaient detectés surtout comme seules infections des plantes apparemment saines. Le SPCSV (70. 6%) avait présenté une incidence élevée, suivi de SPFMV et SPCSV dont le niveau d'infection était le même (8.3%). Basé sur la survie des pousses et l'efficacité de l'élimination de virus, les meilleurs résultats étaient obtenus en exposant les plantules à un regime de température de 32 oC pendant 8 heures sous obscurité et 36 oC pendant 16 heures sous lumière durant quatre semaines. La culture du bout du méristème combinée à la thermothérapie a perimis l'élimination de SPFMV et SPMMV dans 77 % des plants qui étaient au départ infectés avec des virus respectifs. Par ailleurs, l'élimination de SPCSV avait échoué. Mots Cles: Multiplex RT-PCR, thermothérapie, culture de tissue INTRODUCTION Sweetpotato ranks seventh among the most important food crop on fresh weight basis in the world and fifth in over 50 developing countries after rice, wheat, maize and cassava (CIP, 1999). Among the major starch staple crops, it has the largest productivity (production per unit area) (Woolfe, 1992). Annual global production of sweetpotato is greater than 127 million metric tonnes (FAO, 2004). Globally, the bulk of the crop (82.7%) is grown in China (FAO, 2004). In Africa, sweetpotato production is concentrated in countries around Lake Victoria, where it is the second most important food crop after cassava (CIP, 1999). Sweetpotato production is greatly constrained, particularly by viral diseases that cause yield reduction of over 50% (Gibson et al., 1998; Mukasa et al., 2006). In Uganda, sweet potato virus disease (SPVD), caused by co-infection of SPFMV and SPCSV (Gibson et al., 1998), is the most devastating viral disease complex of sweetpotato. Moreover, sweetpotato viruses are mainly spread through healthy-looking vines, which farmers collect from the previous crop for the next cropping cycle. Some reports indicate that these vines might contain at least one virus (Mukasa et al., 2004). The occurrence of single virus infections may make the use of farmer-saved vines unsustainable, especially in areas where vectors are prevalent. This is because singly infected vines can act as source of inoculums, and through vector transmissions lead to mixed infections of different viruses. Dual infections, especially of SPCSV and SPFMV have been reported to interact synergistically resulting in the severe SPVD and yield losses (Gibson et al., 1998; Mukasa et al., 2006). Under field conditions, sweetpotato often becomes infected with up to three different viruses and in a few cases four viruses resulting in decreased quality of planting material (Mukasa et al., 2004). The detection and identification of sweetpotato viruses remain a difficult procedure (Tairo, 2006) and this is complicated by frequent occurrence of mixed infections and synergistic complexes for instance SPVD (Gibson et al., 1998). Serological and PCR procedures have been developed to detect viruses from sweetpotato vines (Gibbs and Padovan, 1993; Colinet et al., 1996; Alicai et al., 1999; Kreuze et al., 2000). However, these procedures were designed to test for a single virus per reaction and this increases the amount of reagents, time and labour required to test each sample. Nonetheless, multiplex reverse transcriptase polymerase chain reaction (RT-PCR) procedure which allows detection of different sweetpotato viruses in a single reaction is still lacking. Control of viral diseases is still difficult in subsistence cropping systems. Targeting insect vectors may not be economically feasible given the subsistence nature of sweetpotato cultivation (Karyeija et al., 1998) and there is insufficient supply of virus-free planting material in tropical Africa to the farmers. The production of disease-free planting material is a major step in the control of sweetpotato viral diseases (CIP, 1999). In Uganda, inadequacy of production, multiplication and distribution of certified virus-free planting material is the major setback in sweetpotato production. SPVD has been associated with the abandonment and extinction of some of the once elite cultivars since severely affected vines cannot be used as sources of planting materials. The production of virus-free planting material could lead to improved quality of seed, and raises the possibility of improving sweetpotato production in Uganda. The objective of this study was to provide an assessment of the incidence of viruses in healthy looking sweetpotato in Uganda and to optimise modern technologies for virus diagnosis and ultimately to develop in vitro techniques for virus elimination for use in clean seed technology and breeding programmes. MATERIALS AND METHODS Plant material. A total of 180 healthy-looking sweetpotato plants were collected from farmers' fields and from a germplasm collection at National Crops Resource and Research Institute (NaCRRI) in Uganda. They were established and kept in a screenhouse at Makerere University Agricultural Research Institute, Kabanyolo (MUARIK). The screenhouse-grown vines were subsequently used in serological and molecular diagnosis for sweetpotato viruses. Four sweetpotato cultivars, namely, Tanzania, New Kawogo, Busia and CIP clone 199004.2 were then used in the virus elimination study. Cultivars Tanzania and New Kawogo are ranked among the popular and widely grown cultivars (Mwanga et al., 1995). Cultivar Busia was selected because it displayed recovery from sweetpotato severe mosaic disease (SPSMD), caused by co-infection of SPCSV and SPMMV (Mukasa et al., 2006). CIP clone 199004.2 is one of the newly introduced orange fleshed cultivars in Uganda. It displays good agronomic traits and probably presents a different genetic background. Healthy in vitro plantlets of clone 199004.2 were obtained from CIP at Lima in Peru. Incidence of viruses in healthy-looking sweetpotato vines. Scions from the 180 sweetpotato plants were grafted to Ipomoea setosa, a sensitive indicator plant for sweetpotato viruses (Schaefers and Terry, 1976). The grafted I. setosa plants were observed for symptoms five weeks post-grafting. Detection of SPCSV, SPFMV, SPMMV and SPCFV was also carried out using nitrocellulose membrane linked immunosorbent assay (NCM-ELISA) as described by Gibbs and Padovan (1993). Each assay for testing the viruses included a positive control provided with the kit from the International Potato Center (CIP). RT-PCR was used to verify the results of plants which tested negative or positive for all the viruses using virus specific primers (Table 1). First strand synthesis of viral cDNA was carried out using random primers and Moloney murine leukaemia virus reverse transcriptase (M-MLV-RT) (Promega, Madison, WI) as described by Nolasco et al. (1993). The PCR products were separated by electrophoresis on 1% agrose gel that contained ethidium bromide. The products were visualised under ultra violet (UV) light for presence or absence of the expected PCR amplicon. Virus isolates and Multiplex RT-PCR. The individual virus isolates, namely, SPMMV-Tor (Mukasa et al., 2003), SPFMV-Nam1 (Kreuze et al., 2000), and SPCSV-Ug (Kreuze et al., 2000) were used. Plants of cv. Tanzania infected with known single viruses were top-grafted with each other to create all possible combinations of the three viruses (SPCSV, SPFMV and SPMMV) commonly found in Uganda. The various infection combinations were used in the subsequent multiplex RT-PCR optimisation and detection assay. Total RNA was extracted from leaf tissue using TRIzol LS Reagent (Invitrogen Ltd, Groningen, Netherlands) according to the manufacturer's instructions. RNA was quantified and tested for quality using a spectrophotometer. An average of 500 µg of RNA per gramme of leaf tissue was used in cDNA synthesis. First strand synthesis of viral cDNA was carried out using random primers and M-MLV-RT (Promega, Madison, WI) as described by Nolasco et al. (1993). Specific primers, namely, CP1 and CP2 (SPCSV); PVD-1 and 10820R (SPFMV) and, PVD-1 and MMD2 (SPMMV), were used in multiplex PCR reaction. PCR amplification was done using the forward and respective reverse primers (Table 1), and the viral cDNA as a template. Several parameters such as magnesium chloride concentration (1.5-2.5 mM), concentration of cDNA (3-5 µl), elongation times (from 1-2 minutes) and annealing temperature were analysed to establish the best conditions for the simultaneous amplification of the three important sweetpotato viruses. The optimised PCR mix consisted of 1 x PCR buffer, 2.5 mM MgCl2, 0.2mM dNTPs, and 0.4 µM forward and reverse primers, 5 µl of sample cDNA, 0.05U µl-1 DyNazyme II (FINNZYMES, Finland) and sterilised distilled and de-ionised water to a final volume of 25 µl was used. The cDNA was denatured at 94 oC for one minute and then amplified through 34 thermal cycles of 94 oC for one minute, 50 oC for 30 seconds, 72 oC for two minutes and ending by a final extension step at 72 oC for 10 minutes. The amplified products were separated by electrophoresis on 1% agrose gel in Tris borate-EDTA (TBE) buffer, stained with ethidium bromide and visualised under UV light. Optimisation of media for micro-propagation. Sweetpotato shoots having about 12 nodes were washed with liquid soap plus distilled water. They were then rinsed in distilled water for 15 minutes, rinsed in 70% alcohol for two minutes, and sterilised with 10% commercial jik (NaOCl) for 15 minutes. They were further rinsed in 5% NaOCl for 10 minutes and then rinsed three times with sterile distilled water. The sterile explants were dissected into single nodes of 0.5-1.0 mm, and placed separately in culture tubes containing 10 ml of MS media. MS media (Murashige and Skoog, 1962) consisted of glycine 2 mg l-1, myo-inositol 100 mg l-1, thiamine-HCL 0.5 mg l-1, nicotinic acid 0.5 mg l-1, pyridoxine-HCL 0.5 mg l-1, sucrose 30 g l-1 and 2.3 g l-1 phytagel. Auxins and cytokinins were added to the MS medium in different concentrations and combinations (Table 2) in order to establish their optimum levels; and 0.1 mg l-1 gibberellic acid (GA3) was also added to all treatment combinations. The pH of the medium was adjusted to 5.7 ± 0.1. The culture medium was autoclaved at 121 oC for 20 minutes and then maintained at 25.0 ± 2.0 oC. The sweetpotato explants were maintained in the culture rooms at 25.0 ± 2.0 oC, 70% relative humidity and illuminated for a 16 hr period using fluorescent lights. In vitro thermotherapy techniques for virus elimination. The in vitro plants infected with SPCSV, SPFMV, SPMMV, and combinations of SPCSV + SPFMV and SPCSV + SPMMV were used as experimental materials. The plantlets were exposed to three diurnal temperature regimes of (i) 28 oC for 8 hr darkness and 32 oC for 16 hr light; (ii) 32 oC for 8 hr darkness, and 36 oC for 16 hr light, and (iii) 34 oC for 8 hr darkness and 40 oC for 16 hr light for four weeks. Each cultivar was subjected to the above temperature regimes. Meristem excision was done to all the recovered plantlets after four weeks of in vitro thermotherapy. Apical meristems of 0.5-1.0 mm, with one to two leaf primodia were excised. Meristems were grown on MS media and were maintained under a 16-hour photoperiod in a culture room. After five weeks of culture, the regenerated plantlets were weaned and potted in a substrate of lake sand:loam soil: saw dust in a ratio of 1:1:1. Acclimatisation in the screenhouse was done in two phases, first under high humidity in a growth chamber for three weeks, and then in open screenhouse conditions. Virus indexing was done using NCM-ELISA, and RT-PCR. RT-PCR was used to verify indexing in the plants which had shown results that were difficult to interpret using NCM-ELISA as described above. Experimental design and statistical analysis. Determination of the best medium for sweetpotato regeneration was carried out as a factorial experiment. Factor A was sweetpotato cultivars; factor B was different hormone combination levels (auxin levels 0, 0.5 and 1.0 mg l-1 and cytokinin levels 0.25 and 0.5 mg l-1). The experiment was replicated three times. In the second experiment, the best temperature regime for sweetpotato virus elimination was determined using plants infected with different initial infecting virus (SPFMV or SPVD). The factors were sweetpotato cultivars (A), and temperature regimes (B). All experimental data were subjected to analysis of variance using GenStat (Lawes Agricultural Trust, 2005) and means were compared using the Least Significance Difference procedure at 5% level of significance. RESULTS Incidence of sweetpotato viruses in healthy-looking plants. Plant infectivity assay using I. setosa indicated that most (87%) of the healthy-looking vines from farmers' fields were infected by at least one virus (Fig. 1). Four viruses namely SPCSV, SPFMV, SPMMV and SPCFV were detected singly or in various combinations in the healthy looking plants. These results were verified by RT-PCR using the virus specific primers (Fig. 2). None of the assayed viruses was detected in the healthy sweetpotato (virus free) samples included in each assay as control. SPCSV (70. 6%) recorded highest incidence, followed by co-infection of SPFMV and SPCSV (8.3%) (Fig. 1). Combinations of SPCSV with SPMMV, and SPCSV with SPCFV were observed in low frequency. Single infections of SPCFV and SPMMV were also observed. However, 26.2% of the plants did not react with any antisera used for the four viruses. Detection of sweetpotato viruses by multiplex RT-PCR. For multiplex RT-PCR optimisation for detection of SPCSV, SPFMV and SPMMV, the amount of MgCl2, amount of cDNA, PCR elongation times and annealing temperatures were established. Specific amplification products with comparable intensities were obtained at 2.5 mM MgCl2 (Fig. 2). The cDNA template quantities of 5 µl per 25 µl reaction mixture showed better PCR products compared to lower quantities of 14 µl. Below 5 µl, some of the PCR products decreased. Increasing the elongation time to 2 minutes in the multiplex RT-PCR increased the amount of longer products (SPFMV; 1.8kb). Although some individual fragments could be specifically amplified at 48 °C (SPCSV and SPMMV) and 58 °C (SPFMV), the annealing temperature of 50 °C was required for the same fragment loci to be co-amplified in multiplex mixtures. Multiplex PCR products of the expected sizes (Table 1) were obtained. Three bands of different molecular weights corresponding to the different viral targets 1800 bp for SPFMV, 1400 bp for SPMMV and 1200 bp for SPCSV were observed in the infected plant samples (Fig. 2). Micropropagation of sweetpotato. Using MS media, micropropagation of sweetpotato the cultivars used did not significantly vary with respect to number of leaves or number of internodes. Differences were observed for shoot height. Cv. New Kawogo significantly attained greater shoot length (3.12 cm) than the rest of the cultivars. Addition of various plant growth regulators to the culture medium significantly (P<0.05) influenced shoot length, number of leaves and internodes at 5 weeks in culture (Table 3). Shoot tip proliferation was evident a week after inoculation on the media under treatment 1 and 4. Treatments 2, 3, 5 and 6 did not support shoot proliferation; the explants developed into callus. In general, cv. Busia (Kabode) took longer to develop well-formed plantlets on media. Plantlets in treatment 1 (0.25 mg l-1 BAP and 0.10 mg l-1 GA3) and treatment 4 (0.50 mg l-1 BAP and 0.10 mg l-1 GA3) had green, healthy shoots. However, a few plantlets showed slight chlorosis after five weeks of culture. The performance of treatment 4 was greater than that of treatment 1 by 45.5, 28.4 and 35.1% with respect to shoot height, number of leaves and number of internodes, respectively. Shoot regeneration from sweetpotato was obtained when plants were cultured in media supplemented with 0.50 mg l-1 of BAP and 0.10 mg l-1 GA3 (Table 4). On contrary, callus was observed in explants cultured on media with 2, 4-D. The auxin was effective in inducing a great amount of soft and loose callus, but it did not induce roots or shoots of sweetpotato. Callus production was more vigorous in the presence of higher 2, 4-D concentrations and remained fresh for a period of about four weeks before degeneration. Virus elimination by in vitro thermotherapy. There were significant differences between cultivars (Table 4) and temperature regimes (P<0.05) (Table 5) on virus elimination efficiency. The thermotherapy treatment given to in vitro plantlets enhanced SPFMV and SPMMV eradication. Best results of thermotherapy on virus elimination and plantlet recovery were obtained with daily temperature regimes of 32 oC for 8 hr darkness and 36 oC for 16 hr light. This temperature regime proved to be the most efficient in eliminating viruses in a high number of plantlets as reflected by the high percentage of recovery (97.2%) (Table 5). The elimination of SPFMV was genotype dependent in single infections (Table 4). All meristem derived plants of cv. New Kawogo were SPFMV-free, while Tanzania and CIP clone 199004.2 had some plants which were not virus free in the medium and low temperature regimes. All plants which were exposed to the high temperature regime were free of SPFMV in the three cultivars (Table 5). Nonetheless, the percentage of shoots freed from SPFMV in single infections was high (87%) compared to SPFMV in double infections with SPCSV (73%). There was significantly (P<0.05) higher virus elimination efficiency obtained from the third temperature regime, though this led to low plantlet recovery. Shoot cultures generally showed heat stress ranging from leaf chlorosis to wilting. Leaf and shoot tips turned brown and necrotic. Survival of plantlets from heat stress varied among the cultivars. New Kawogo (73.4%) had high plantlet recovery at temperature regime of 34 oC for 8 hr darkness, and 40 oC for 16 hr light. Heat therapy at 32 oC for 8 hr darkness and 36 oC for 16 hr light, resulted in more plants surviving heat stress than at a higher temperatures. Again New Kawogo (83.4%) had high plantlet survival, followed, by Tanzania (76.7%), CIP clone 199004.2 (73.4%), and Busia (60%). All plantlets of cv. New Kawogo survived thermotherapy of 28 oC for 8 hr darkness, and 32 oC for 16 hr light. The other cultivars also had high plantlet survival in the low temperature regimes ranging from 60 to 90%. Cultivar Busia was the most affected by in vitro thermotherapy in the three temperature regimes. After meristem excision, the recovery of the plantlets which had been subjected to heat treatment was high (69-97%) for the four cultivars. The efficiency of regeneration ranged from 70 to 82% (Table 4) for all cultivars regardless of the temperature regime exposed. There was no observable somaclonal variation as all the established plants of the same cultivar looked morphologically similar to the source plant. Overall, based on shoot survival and effectiveness of virus elimination, the best results were obtained by exposing plantlets to daily temperature regime of 32 oC for 8 hr darkness, and 36 oC for 16 hr light for four weeks. DISCUSSION Incidence of sweetpotato viruses in healthy-looking plants. The high incidence of single virus infections in health-looking vines may suggest a number of implications. First, this observation emphasises the fact that use of farmer saved vines contributes to perpetuation of the individual viruses. This may particularly the case for SPCSV and SPFMV that were observed in higher proportions compared to SPMMV and SPCFV (Figure 1). Secondly, the perpetuation of these single viruses from one crop to the next means that there is always available inoculum from which dual and synergistic interactions could be reconstituted. Co-infection of SPCSV with SPFMV results in the severe SPVD and during this synergistic interaction the titres of SPFMV are elevated by over 600 folds while the titres of SPCSV remain unchanged (Karyeija et al., 2000a; Mukasa et al., 2006).This implies that, the farmers' practice of selecting healthy-looking vines as the source of planting material would to minimise keep SPFMV inoculums low since farmers would select against SPVD affected vines that harbor high tires of SPFMV. Among the sampled health looking vines only 8.3% were infected by SPVD. These plants could have been collected at an early stage in the development of SPVD before manifestation of the obvious and severe symptoms. It also suggests that while selecting for health-looking vines that are mostly infected by single viruses, it possible to select dually infected vines. The high incidence of SPCSV in healthy looking sweetpotato (Fig. 1) could be explained by the abundance of whiteflies which are the vectors of SPCSV in sweetpotato fields. Field observations at MUARIK indicated that each plant could be infested by over 200 whiteflies and this increases the likelihood of being infected with SPCSV. However, aphids, the vectors of SPFMV rarely colonise the sweetpotato plants in Uganda (Gibson et al., 1997). This could explain why SPFMV is less common in the healthy looking vines. The fact that SPCSV was the most common virus in the healthy looking vines suggests that SPFMV (or SPMMV) is the limiting virus to complete the SPVD (or SPSMD) development. However, our results contrast with the incidence of sweetpotato viruses in wild Ipomoea species where SPFMV is reportedly more prevalent than SPCSV (Tugume et al., 2007). Very few (1.6%) plants had SPCFV in single or mixed infection (Fig. 1) probably because SPCFV is also transmitted by aphids which are rarely found on sweetpotato (Aritua et al., 1998). It is clear from this study that without a planned virus eradication system for production of virus free sweetpotato planting material, farmers may not obtain a reasonable proportion of virus-free planting material by selecting for healthy looking vines for the next cropping cycle. Detection of sweetpotato viruses by Multiplex RT-PCR. It is clear from this study that multiplex RT-PCR can reliably be used to simultaneous detect SPCSV, SPFMV and SPMMV in a single step and tube reaction (Fig. 2). Hitherto, separate RT-PCR tests were required for the reliable detection of the three viruses, which greatly increased the amount of reagents, time, labour and cost required to test each sample. Multiplex RT-PCR assay is often cheaper, easier and quicker than for monospecific RT-PCR (Bertolini et al., 2001). This is the first report describing the use of three primer multiplex set for one step RT-PCR in a single tube for sweetpotato RNA viruses. The procedure could thus be used as a common method in the detection of viruses, especially those that lack commercially available antibodies. Presently, sweetpotato virus antibodies are only supplied by the International Potato Centre (CIP; Lima, Peru) to many developing countries in small diagnostic quantities. This limits utilisation of ELISA for example by large-scale seed certification programmes in developing countries. Micropropagation of sweetpotato. The results obtained with one genotype in a given medium differed from those obtained with other genotypes (Table 3). This could be explained by the differences in the growth regulators used for the establishment and initiation of in vitro cultures of sweetpotato cultivars (Tanzania, New Kawogo, Busia and CIP clone 199004.2). The results showed that BAP was crucial for stimulating shoot growth and development. Moreover, the concentration of this cytokinin strongly influenced shoot length of all cultivars. Research on various plant regeneration systems indicate that morphogenesis is regulated by an intricate balance of endogenous auxin and cytokinin (Hutchinson et al., 1996). The addition of 0.50 mg l-1 of BAP and 0.10 mg l-1 GA3 to the propagation medium was essential to stimulate sprouting, growth and lengthening of the shoots in culture. However, the inclusion of auxins in the culture media was not necessary for the establishment of shoot tip cultures since it resulted in the formation of unwanted calli. A possible explanation for this might be that the explants would act as active centres of auxin biosynthesis. Virus elimination by in vitro thermotherapy. The thermotherapy treatment given to in vitro plantlets enhanced SPFMV and SPMMV eradication (Table 5) but at lower temperature regimes, lower recovery levels were recorded indicating higher recovery with increasing temperature (Table 4). These results are not surprising since thermotherapy given to in vitro plants prior to meristem excision has been found to give fewer virus-infected meristem-cultured plants in various vegetatively propagated species (Acedo, 2006) and specifically in sweetpotato (Green and Lo, 1989; CIP, 1999). However, different cultivars have also been reported to behave differently when subjected to in vitro conditions and high temperature regimes, a situation that had not been tested for Ugandan sweetpotato germplasm. The results indicate differences in elimination efficiency of RNA viruses from sweetpotato (Table 4). This could be attributed to differences in the infecting virus and cultivar response to the infection. SPFMV was easily eliminated when present as a single virus in the plantlets, but was slightly difficult when present in mixed infections. This could be attributed to the occurrence of SPFMV in almost all tissue because of the increase in its titres due to the presence of SPCSV. Karyeija (2000a,b) reported that titres and distribution of potyviruses (SPFMV and SPMMV) in the infected plants are enhanced in the presence of SPCSV due to the synergistic interaction. Thus, it would be expected to be more difficult to eliminate viruses mixed infections than single infections. However, the percentage recovery of SPFMV or SPMMV freed plants was high for all cultivars in mixed infection. Notably the elimination of SPFMV was higher in mixed infections compared to SPMMV. This could have been due to cultivar effects since elimination of the two viruses were not from the same genotype. The success of in vitro thermotherapy on virus elimination in sweetpotato appear to be due to the combined effects of various factors, including the use of micropropagated shoots of the diseased cultivars instead of pot grown plants as a source of meristems, the effective heat treatment and the technique of meristem isolation. Kassanis (1954) concluded that increased temperatures create disadvantageous conditions for virus replication due to competition among the rapidly diving host cells and virus particles. Thus, temperatures higher than 32 oC may have created an altered balance between synthesis and degradation of the virions causing the elimination of viruses. Mink et al. (1998) later suggested that increased temperatures destroy essential chemical processes in the viral life cycle. Meristem culture per se could also have contributed to elimination of sweetpotato viruses in this study due to uneven distribution of viruses in the plant tissues. This technique takes advantage of the fact that some viruses are unable to colonise meristem regions because of inhibition of replication and restriction of their movement (Faccioli and Moran, 1998). The virus may fail to keep up with the pace of rapidly dividing cells at the growing point. This may render the virus unable to move across the plant cells through plasmodesmata to the meristem and the high concentration of auxin within the meristem inhibits the virus replication. ACKNOWLEDGEMENT Materials used in this study were assembled with the support from International Foundation for Science (IFS, grant No. C/3569). We also gratefully acknowledge the Regional Universities Forum for capacity building (2005 RU CG 005) and the East African Regional Network for Biotechnology, Biosafety and Biotechnology Policy Development Programme (BIO-EARN Cassava/Sweetpotato Project 02) for the financial support towards the tissue culture and molecular studies, respectively. REFERENCES
Copyright 2010 - African Crop Science Journal The following images related to this document are available:Photo images[cs10026t3.jpg] [cs10026t2.jpg] [cs10026f1.jpg] [cs10026f2.jpg] [cs10026t5.jpg] [cs10026t1.jpg] [cs10026t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}