 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
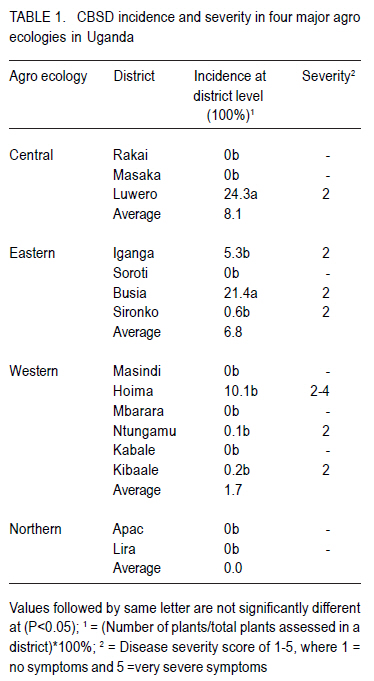
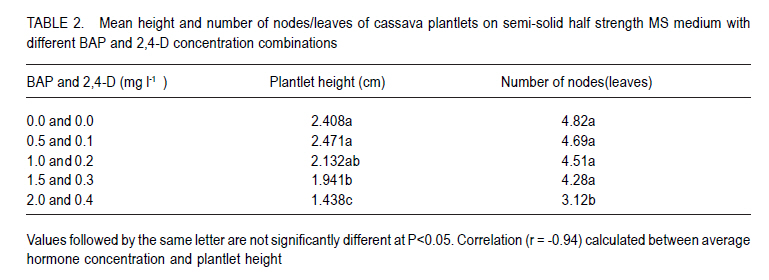
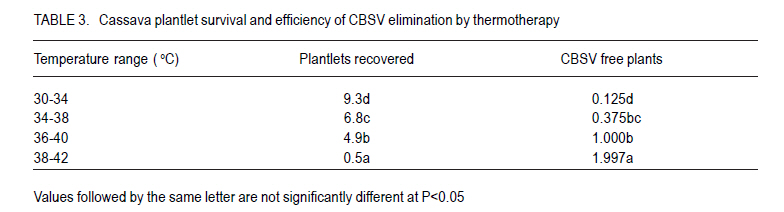
African Crop Science Journal, Vol. 18, No. 3, 2010, pp. 235 - 241 Optimisation of in vitro techniques for Cassava brown streak virus elimination from infected cassava clones P. Wasswa, T. Alicai1 and S.B. Mukasa Department of Crop Science, Makerere University, P. O. Box
7062, Kampala, Uganda
Corresponding author: sbmukasa@agric.mak.ac.ug Code Number: cs10027 Abstract Cassava brown streak disease (CBSD), caused by Cassava brown streak virus (CBSV), is an economically important disease of cassava (Manihot esculenta Crantz) in East Africa. The objective of this study was to optimise in vitro techniques for CBSV elimination from infected Ugandan cassava cultivars. Using semi-solid half-strength Murashige and Skoog (MS) basal medium, hormone concentration and heat treatment regimes were optimised for micropropagation of farmer's preferred cassava cultivars and CBSV elimination. Single nodes from young cassava stems were cultured for four weeks on ½MS medium supplemented with 6-benzyl amino purine (BAP) and 2,4-dichlorophenoxy acetic acid (2,4-D). The BAP and 2,4-D concentration (mg l-1) combinations used were 0.0 and 0.0, 0.5 and 0.1, 1.0 and 0.2, 1.5 and 0.3, and 2.0 and 0.4, respectively. The optimum medium was used for in vitro thermotherapy using four temperature regimes, namely 30-34, 34-38, 36-40 and 38-42 oC for 8 hours darkness and 16 hours light, respectively, for four weeks. The best plantlet growth in terms of height was observed on MS medium supplemented with 0.5 mg l-1 BAP and 0.1 mg l-1 2,4-D. Highest CBSV elimination efficiency of 40%, with 49% plantlet survival was observed at 36 oC for 8 hours darkness and 40 oC for 16 hours light. These results indicate that in vitro techniques can greatly enhance CBSV elimination and, thus, provide a means of CBSD management through dissemination and conservation of popular but CBSD susceptible cultivars. Key Words: Manihot esculenta, Murashige and Skoog medium, thermotherapy RÉSUMÉ La maladie de la striure brune du manioc (CBSD) causée par le virus de la striure brune (CBSV), est une maladie économiquement importante du manioc (Manihot esculenta Crantz) en Afrique de l'Est. L'objectif de cette étude était d'optimiser les techniques in vitro pour l'élimination du CBSV des cultivars ougandais infectés. En utilisant le médium basal semi-solid half strength Murashige and Skoog, la concentration d'hormone et les regimes de traitement de chaleur étaient optimisés pour la micropropagation des cultivars de manioc préférés par les fermiers et l'élimination du CBSV. Les nodes simples des jeunes tiges de manioc étaient cultivés pendant quatre semaines sur le médium ½ MS supplémentés avec 6-benzyl amino purine (BAP) et 2,4-dichlorophenoxy acetic acid. Les combinaisons de concentrations (mg l-1) de BAP et de 2.4-D utilisées étaient 0.0 et 0.0, 0.5 et 0.1, 1.0 et 0.2, 1.5 et 0.3, et 2.0 et 0.4, respectivement. Le médium optimum était utilisé pour la thermothérapie utilisant quatre regimes de température dont 30-34, 34-38, 36-40 et 38-42 oC pendant 8 heures en obscurité et 16 heures sous lumière, respectivement pendant quatre semaines. La meilleure croissance en hauteur des plantules était observée sur le médium MS supplémenté avec 0.5 mg l-1 BAP et 0.1 mg l-1 2,4-D. L'efficience la plus élevée de 40 % en terme d'élimination du CBSV, avec 49 % de la survie de plantules était observée à 36 oC pendant 8 heures sous obscurité et 40 oC pendant 16 heures sous lumière. Les résultats indiquent que les techniques in vitro peuvent grandement promouvoir l'élimination du CBSV et offrent ainsi un moyen de gestion de CBSD à travers la dissemination et la conservation des cultivars aussi populaires que susceptibles au SBSD. Mots Cles: Manihot esculenta, Murashige and Skoog medium, thermothérapie INTRODUCTION Cassava brown streak disease (CBSD) is a destructive systemic disease of cassava (Manihot esculenta Crantz) caused by Cassava brown streak virus (CBSV) (genus Ipomovirus, family Potyviridae) (Monger et al., 2001). The disease damages leaves, stems and roots that often become necrotic and constricted, thus, reducing crop yield and quality. The affected roots are not fit for food or feed (Hillocks et al., 2001). CBSD was once eliminated from Uganda by rouging when it had been inadvertently introduced in planting material from Tanzania (Jameson, 1964). However, recent reports show re-emergency of the disease (Alicai et al., 2007) and several farmer-preferred cassava cultivars are seriously affected. Cultivars such as TME 14, TME 204, 2691 and Akena have been bred and selected for yield, quality and resistance to African cassava mosaic disease (ACMD) caused by African cassava mosaic virus (ACMV) (genus Begomovirus, family Germiniviridae). But all these cultivars have proved to be very susceptible to CBSD (Alicai et al., 2007). Biotechnological techniques such as in vitro thermotherapy and meristem tip culture can be used for virus elimination and regeneration of plantlets from tissues where viruses have been excluded (Zapata et al., 1995). Apical meristems in the infected plants are generally either free or carry very low titre of the virus. The explanations for the escape of the meristem by virus invasion are (i) some viruses move readily in a plant body through the vascular system which is absent in meristem, (ii) high metabolic activity in the actively diving meristem cell does not allow virus replication due to competition, and (iii) a high endogenous auxin level in shoot apices may also inhibit virus multiplication. During the heat treatment, the high temperature is unsuitable for virus replication which lowers virus titre and enhances elimination of viruses from infected plants (Walkey, 1976). The success of virus elimination depends on the plant virus species, the host (cultivar), and whether the plant is single or mixed infected (Allam, 2000). Elevated temperatures in vitro have been effective in eliminating several viruses known to infect vegetatively propagated crops (Allam, 2000). These methods and techniques, however, had not been optimised for the Ugandan cassava cultivars. Thus, the aim of the study was to optimise in vitro techniques for CBSV elimination from infected cassava cultivars in Uganda. MATERIALS AND METHODS Cassava material. Stems of 133 popularly grown cassava cultivars and/or apparently CBSV infected cassava plants were collected from 15 districts in Uganda (Table 1). The districts represented agro-ecologies where high CBSD prevalence had been reported and cassava is popularly grown. Districts were also selected from high altitude areas where CBSD occurrence was previously reported to be uncommon (Alicai et al., 2007). In each district, ten cassava fields each of at least 0.5 hectare ranging from 6 months to 2 years old were selected along major and feeder roads traversing the district. The distance between sampled fields was 6-7 km. Within each field, 30 cassava plants were chosen along transects of a field and assessed for CBSD incidence (Madden and Hughens, 1999) and severity (Gondwe et al., 2002). CBSD incidence data were subjected to analysis of variance (ANOVA) using SPSS 11.0 for Windows at P<0.05. Three stem cuttings were collected per cultivar and established in a screenhouse at Makerere University Agricultural Research Institute, Kabanyolo (MUARIK). Sprouted cassava cuttings were diagnosed to confirm CBSV infection using reverse transcription-polymerase chain reaction (RT-PCR) technique with virus specific primers (Monger et al., 2001). Optimising MS medium for cassava micropropagation. Ten single node cuttings from young stems of each of the ten popularly grown cassava cultivars were sterlised in 15% jik (0.00525% w/v sodium hypochlorite) for 15 minutes, then in 70% ethanol for 5 minutes and finally rinsed four times in double distilled water. The nodes were cultured on semi-solid half-strength Murashige and Skoog (MS) basal medium (Murashige and Skoog, 1962), supplemented with BAP and 2,4-D concentration (mg l-1) combinations of 0.0 and 0.0, 0.5 and 0.1, 1.0 and 0.2, 1.5 and 0.3, and 2.0 and 0.4, respectively. The cultures were incubated under white fluorescent light with a 16 hour photoperiod at 25±1 oC. Then shoot length, number of nodes (leaves) and root development were recorded every week for four weeks and, thereafter subcultured. The optimum medium was used for subsequent meristem tip culture and thermotherapy techniques for CBSV elimination. The experiment was laid in a completely randomised design (CRD) and each medium treatment was replicated three times. Shoot length and number of leaves (nodes) data were subjected to ANOVA using SPSS 11.0 for Windows. Where the ANOVA indicated significant (P<0.05) difference, the means were separated using Student-Newman-Keuls (SNK). The correlation between plantlet height and hormone concentration was calculated. Optimising in vitro thermotherapy for CBSV elimination. Ten CBSV infected in vitro cassava plantlets were exposed to diurnal temperature regimes of 30 and 34, 34 and 38, 36 and 40, and 38 and 42 oC for 8 hours of darkness and 16 hours of light, respectively, for four weeks. Then plantlet survival was recorded. Meristem tips (~0.5 mm) were excised and cultured on the optimum medium (MS medium supplemented with 0.5 mg l-1 BAP and 0.1 mg l-1 2,4-D) for four weeks. Plantlets were weaned in a growth chamber under 80% humidity conditions for eight weeks and thereafter in an insect proof screenhouse conditions for six weeks. Plantlets were then indexed for CBSV using RT-PCR to establish CBSV elimination efficiency. The experiment was laid in completely randomised design and was replicated three times for each temperature regime for each of the ten cultivars. Each cassava sample was scored as a binary variable (virus positive = 1 and virus negative = 0). Mean plantlet recovery values and the significance of recovering virus free plants from in vitro meristem tip culture and thermotherapy techniques were established by subjecting the data to ANOVA using SPSS 11.0 for Windows. Where the ANOVA indicated significant (P<0.05) difference, the means were separated using SNK. RESULTS CBSD incidence and severity in Uganda. Apparent CBSD symptoms were observed in farmers' fields in 7 out of the 15 districts, but with varying severity levels (Table 1). The most commonly observed symptoms were the yellow blotches on mature leaves and feathery chlorosis closely oriented along secondary and tertiary veins. Root constrictions and corky necrotic lesions were less frequently observed than foliar symptoms. There was no relationship between CBSD incidence and altitude. In the northern region, districts of Apac and Lira, CBSD symptoms were not observed. (Table 1). Other districts that did not record CBSD incidence were Rakai, Masaka, Soroti, Masindi, Mbarara and Kabale (Table 1). Districts that had cassava plants with CBSD symptoms were Luwero, Iganga, Busia, Sironko, Hoima, Ntungamo and Kibaale. CBSD incidence was highest in Luwero district (24.3%) while severity was highest in Hoima district (2-4) (Table 1). In vitro micropropagation of cassava. Cassava cultivars varied widely in response to in vitro culture with some cultivars responding better than others. In general, elongation of axillary buds and emergence of new leaves were observed 10 weeks after initiation of the explants. Root formation was evident 3 weeks after initiation. When subcultured, it took 2 and 4 weeks for shoot and root formation, respectively. There was a strong negative correlation (r = -0.94) between plantlet height and hormone concentration (Table 2). The highest mean plantlet height (2.471 cm) was observed 4 weeks after initiation on semi-solid half-strength MS medium supplemented with 0.5 mg l-1 BAP and 0.1 mg l-1 2,4-D (Table 2). However, this highest plantlet height (2.471 cm) did not significantly (P>0.05) differ from the plantlet height (2.408 cm) observed on MS medium with no hormones (Table 2). Mean node (leaf) number (4.82) per plantlet was highest on medium without BAP and 2,4-D (Table 2). In vitro thermotherapy for CBSV elimination. Cassava plantlets had varying response to heat treatment (Table 3). Heat stress ranged from scotched leaves and shoot tips in tolerant cultivars to total death of susceptible cultivars for a given temperature regime. Generally, low plantlet survival (5%) was observed at high temperature regime (38 - 42 oC), and higher plantlet survival (93%) at lower temperatures (30 - 34 oC) (Table 3). However, thermotherapy increasingly enhanced recovery of CBSV free plants with increase in temperature. Also, CBSV elimination efficiency significantly (P<0.05) varied among temperature regimes (Table 3). Overall, optimum CBSV elimination efficiency of 40% with 49% mean plantlet survival was observed at 36 oC for 8 hours darkness and 40 oC for 16 hours light (Table 3). DISCUSSION CBSD incidence and severity in Uganda. Incidence data in low verses high altitude areas (districts) did not support the fact that whiteflies are the major vehicles of CBSV as previously reckoned by Maruthi et al. (2005) and Alicai et al. (2007). In fact, some low land districts of Rakai, Masaka, Masindi, Mbarara, Apac and Lira did not record any CBSD incidence, yet they habour high incidences of whiteflies (Alicai et al., 2007). The major factor contributing to the high CBSD incidence in central region (Table 1) may, therefore, be attributed to unrestricted movement of planting material between and within regions. The areas with high incidence of CBSD, e.g. Mukono district, have been associated with high human activity regarding exchange of planting materials, where several NGOs activity have played part in introducing new and high yield materials. These materials; e.g. cassava cultivars TME 14 and TME 204 were bred for resistance to ACMD are currently being reported to be very susceptible to CBSD (Alicai et al., 2007). Similarly, the early epidemics of CBSD in Uganda were attributed to introduction of planting material from Tanzania where CBSD is endemic (Jameson, 1964). Alicai et al. (2007) also observed CBSD to be common on the newly introduced cassava cultivars in several districts of Uganda. The fact that farmers obtain cuttings, both exotic and indigenous cassava cultivars, from their previous crop or from their neighbours, suggests that without a deliberate programme for restricting movement of infected material would perpetuate the CBSD spread. The variation in CBSD symptom severity between Hoima and other districts is probably due to the differences between seasons, crop age and cultivars grown. Hillocks et al. (1999) observed that young plants and leaves commonly do not show CBSD symptoms, whereas mature symptomatic leaves soon dry and drop off. Mware et al. (2009), just like us, observed severity variability among cassava cultivars in Kenya, depicting differential cultivar sensitivity to CBSD. They observed that some cultivars had distinct CBSD symptoms, others expressed only mild symptoms, while some had no symptoms. These observations are consistent with earlier work done in Tanzania by Hillocks et al. (2001). Besides environmental conditions, age of crop and type of cultivar, CBSD severity may vary due to the differences between CBSV isolates. Indeed, CBSV isolates differ in the symptoms they induce on indicator plants (Monger et al., 2001). Mbanzibwa et al. (2009) identified two different CBSV isolates infecting cassava cultivars in Uganda and there is an on-going study in the country to see if the two isolates cause different symptoms in infected cassava cultivars. In vitro micropropagation of cassava. The relatively good plantlet height and number of nodes produced in a relatively short period of four weeks (Table 2) suggests a high potential for in vitro micropropagation of cassava. This is consistent with earlier findings from a study done in Philippines by Acedo (2002). He found out that 159,432 to 3,774,873 cassava plantlets could be produced from one nodal explant per year. The problem with in vitro micropropagation techniques is that cultivars vary widely in response to in vitro culture (Kaiser and Teemba, 1979). This makes the use of a general protocol difficult. This is evidenced by the differences in the influence of growth regulators on the growth of in vitro cassava cultivars used in this study (Table 2). The general negative effect of adding hormones observed in the study (Table 2) implies that inclusion of BAP and 2,4-D in the culture media was not essential for in vitro micropropagation of cassava. Actually, supplementation of the medium with these growth hormones instead suppressed cassava organogenesis with the cultivars used. Moreover, best plantlet height observed on MS medium supplemented with 0.5 mg l-1 BAP and 0.1 mg l-1 2,4-D did not significantly (P<0.05) vary from that observed on MS medium with no hormones. This suggests that cassava explants may act as active centers for cytokinin and auxin synthesis. Therefore, enough amounts of the hormones could be produced by the explants, implying that no need for supplementation. This reduces on the cost of production and possible mutations which arise especially from multiple subcultures (Larkin and Scowcroft, 1981). In vitro thermotherapy for CBSV elimination. High plantlet mortality at high temperature could be attributed to high heat stress on plantlets. High temperatures denature enzymes, thus interfering with essential physiological processes of the plant (Kaiser and Teemba, 1979). On the contrary, the observable improved CBSV elimination efficiency at high temperatures suggests that thermotherapy technique is a reliable means for CBSV elimination. During heat treatment, there are probably unsuitable conditions for virus replication and movement, thus the growing tip elongates faster than the rate at which the virus moves into the meristem tip. This greatly increases the meristem tip region that is virus free, thus enhancing recovery of virus-freed plants from infected clones. Similarly, it was reckoned that, a high metabolic activity in the actively dividing meristem cells does not allow virus replication due to competition for resources (Valentine et al., 2001). Allam (2000) suggested that under high temperature, the union of the protein sub-units that protect the nucleic acid of the virus becomes weaker and temporal fissures appear, allowing attack by nucleases. Therefore, the high CBSV elimination efficiency at high temperature could be attributed to the fact that increased temperatures destroy essential chemical processes in the virus life cycle. Presence of inhibiting compounds in the medium such as amino acids, cytokinin and auxins is one other factor that probably contributed to the elimination of CBSV from cassava explants (Paula, 2000). In fact, it has been reported that shoot apices have a high indigenous auxin level which may inhibit virus multiplication (Valentine et al., 2001). And, in general some cells, particularly of the reproductive organs, apical shoot and root meristems might be free from viruses or carry very low virus titers (Mori and Hosokawa, 1977). CONCLUSION In general, it is possible to rapidly micro-propagate cassava plantlets and to achieve CBSV elimination following in vitro culture and thermotherapy techniques. Farmers should therefore be strongly encouraged to use in vitro raised materials that are adequately diagnosed for CBSV. This can reduce the practice of moving cassava stems of apparently healthy cassava between and within regions thus enhancing integrated CBSD management in cassava fields in Uganda. Additional future studies are, however, needed to ascertain the feasibility of large scale production of in vitro cassava in Uganda. ACKNOWLEDGEMENT This study was funded by Swedish International Development Cooperation Agency/SAREC (Sida/SAREC) through the East African Regional Research Network for Biotechnology, Biosaftey and Biotechnology Policy Development (BIO-EARN/02). The contribution of National Cassava Program of the National Crops Resources Research Institute - Namulonge Uganda is highly appreciated. REFERENCES
The following images related to this document are available:Photo images[cs10027t2.jpg] [cs10027t3.jpg] [cs10027t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}