 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
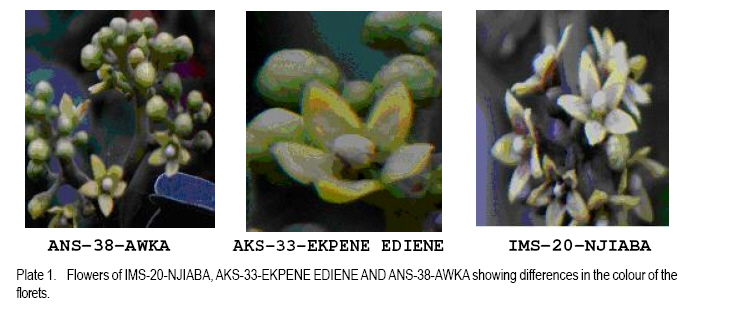
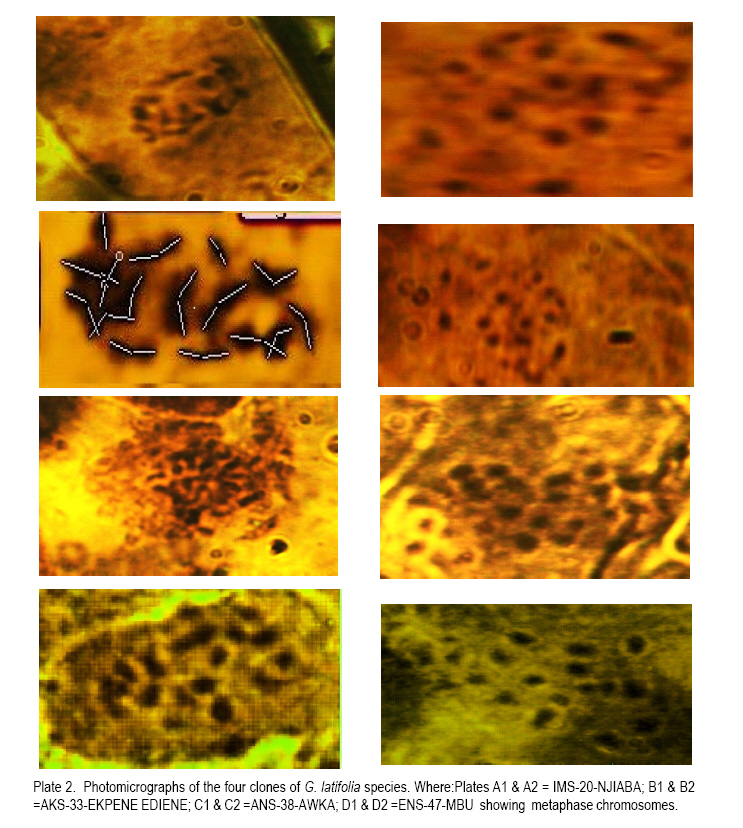
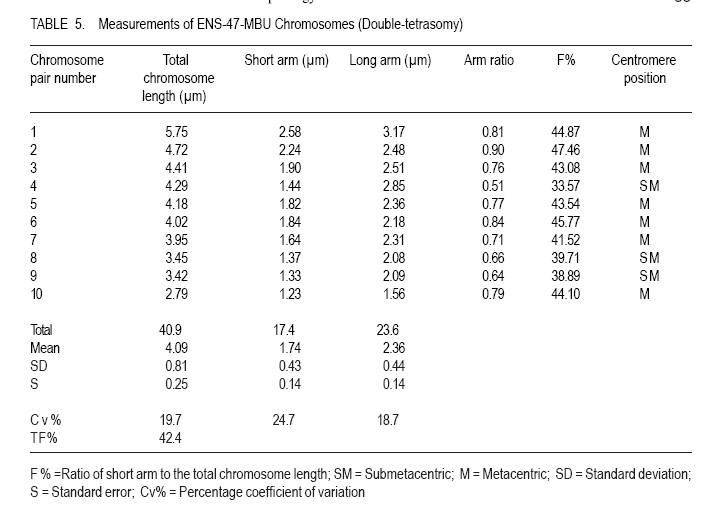
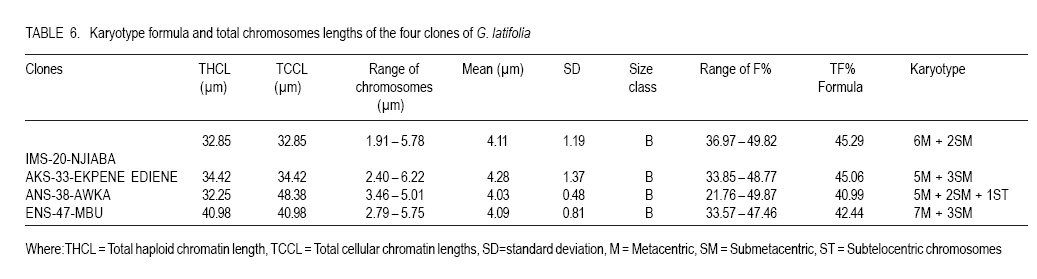
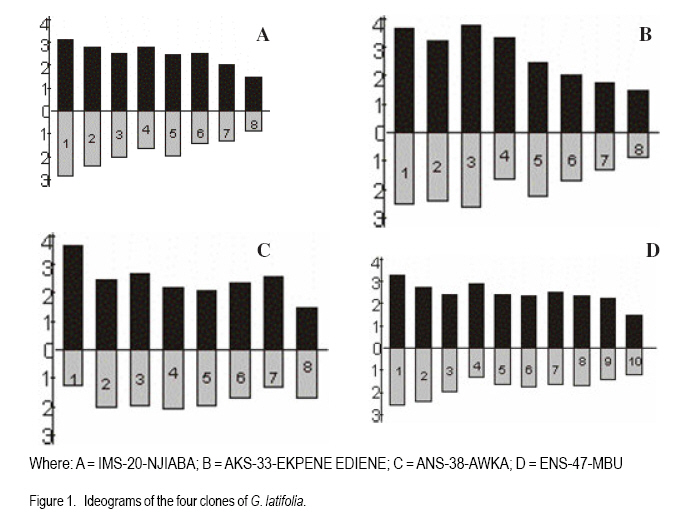
African Crop Science Journal, Vol. 19, No. 1, 2010, pp. 29 - 38 MORPHOLOGY AND CHROMOSOME NUMBERS OF Gongronema latifolia BENTH. CLONES FROM NIGERIA C.U. Agbo and N.U. Ukwu Department of Crop Science, University of Nigeria, Nsukka, Nigeria (Received 20 June, 2010; accepted 23 November, 2010) Code Number: c11003 ABSTRACT Cytological studies on the root-tips of four clones of Gongronema latifolia Benth. were conducted to identify cross-compatible clones for possible improvement through hybridisation. The results showed that the diploid chromosomes number in G. latifolia was 2n = 16. Clones, IMS-20-NJIABA, AKS-33-EKPENE EDIENE, ANS-38-AWKA and ENS-47-MBU used in the study had 2n = 16, 2n = 16, 2n = 24 and 2n = 20 chromosomes, respectively. This showed that ANS-38-AWKA and ENS-47-MBU are polyploids. Chromosomes were metacentric and submetacentric, with only a pair of subtelocentric chromosome in ANS-38-AWKA. Chromosome lengths ranged from 4.03 to 4.28 µm, and were generally of the B-size. The F values ranged from 21.76 to 49.87%. Chromosomal variations among the clones suggest presence of high levels of cross incompatibility between clones that are not of the same ploidy level. Hence, only clones IMS-20-NJIABA and AKS-33-EKPENE EDIENE that are of the same ploidy level will be cross compatible. Key Words: Cross compatibility, metacentric and submetacentric, ploidy RÉSUMÉ Les études cytologiques de racines de quatre clones de Gongronema latifolia Benth, étaient conduits pour identifier des croisements compatibles de clones pour une possible amelioration par hybridation. Les résultats ont montré que le nombre de chromosomes diploïdes dans G. latifolia était de 2n = 16. Les clones utilisés pour cette étude étaient IMS-20-NJIABA, AKS-33-EKPENE EDIENE, ANS-38-AWKA ainsi que ENS-47-MBU et avaient 2n = 16, 2n = 16, 2n = 24 et 2n = 20 chromosomes, respectivement. Ceci a montré que ANS-38-AWKA et ENS-47-MBU sont polyploïdes. Les chromosomes étaient métacentriques et submétacentriques, avec seulement une paire de chromosome sub-télocentrique dans ANS-38-AWKA. La grandeur de chromosomes variait de 4.03 à4.28 µm et étaient généralement de la taille de B. Les valeurs de F variaient de 21.76 à 49.87 %. De variations de chromosomes parmi les clones suggèrent l'existance d'un niveau élevé d'incompatibilité de croisement entre les clones qui ne sont pas dee même niveau de ploïdie. Ainsi, seuls les clones IMS-20-NJIABA et AKS-33- EKPENE EDIENE dont le niveau de ploïdie était le même seront compatiblement croisés. Mot clés: Compatibilité de croisement, métacentrique et sub-métacentrique, ploïdie INTRODUCTION Gongronema latifolia Benth., a non-wood forest product is a leafy vegetable belonging to the order Apocynales, family Asclepiadaceae, and sub-family Asclepiadciopeae (Nielsen, 1965). It is a common forest climber with hollow stems and broadly ovate leaves that are widely cordate at the base. It grows in the forests of Southeastern Nigeria (Akpan, 2004), and is of West African Origin (Nielsen, 1965). It is commonly called "Utazi" in Southeastern Nigeria, and "Arokeke" in Southwestern Nigeria (Ugochukwu et al., 2003). It is a good source of vitamins, minerals and proteins (Okafor, 2005). The medicinal importance of G. latifolia cannot be overemphasized. The plant plays a vital role in the treatment and prevention of varied health related problems including liver diseases, diabetes mellitus, high blood pressure, loss of appetite, dysentery, stomach pains, worm infections, cough and malaria fever (Agbo et al., 2005; Okafor, 2005). Medicinal importance of the plant is further elaborated by the presence of five bioactive compounds including alkaloids, saponins, tannins, flavonoides, and glycosides in leaves, which was suggested to proffer varied pharmacological effects on the species (Gamaniel and Akah, 1996). Reports on the cytology of the plant species are scanty. However, there are a lot of variations in the morphology and chemical composition of the plant species. Morphological variation such as flower colour and leaf sizes could be caused by variations in chromosome numbers, and sizes and shapes. Thus, knowledge of the cytology of the different clones of the plant species can serve as an important guide in the planning of a successful breeding programme (Armstrong, 1987). The objectives of this research were to: (i) determine the chromosome numbers and shape of four different clones of the plant species with distinct morphologies in Nigeria; and (ii) identify cross-compatible clones for possible improvement through hybridisation. MATERIALS AND METHODS This research was conducted in the Cytogenetic Laboratory of the Department of Crop Science, University of Nigeria, Nsukka. Root-tips of Gongronema latifolia of the four clones (IMS-20-NJIABA, AKS-33-EKPENE EDIENE, ANS-38-AWKA and ENS-47-MBU) were obtained from stem cuttings rooted in a mixture of topsoil and sawdust medium in polyethylene bags at 1:1 ratio. The root-tips were harvested between the hours of 810 am. and pretreated in a solution of 8-hydroxyquiniline for one hour in the refrigerator. The pretreated root-tips were rinsed in distilled water and fixed in Carnoy's fluid (70% ethanol + 30% acetic acid) for 24 hours at room temperature. The fixed root-tips were hydrolyzed in 18% Hcl for 3 5 minutes to soften the root-tips for squashing and afterwards, were rinsed thoroughly in distilled water. They were then transferred back to the fixing fluid to enable the chromosome pick up stains. Using a pair of forceps and a dissecting needle, the milky coloured meristematic zone was excised and squashed in 2 drops of F. L. P. O. stain after dissecting. The outer cuticles were removed, and cover slips used to cover the slides as gently as possible as to avoid air bubbles. The slides were labeled and left for 24 hours to allow for proper staining and pressed in the middle of a double folded filter paper to remove excess stain and to flatten the cells for viewing. The contents of the slides were observed at a magnification of x 10 and x 40, respectively. Good cells with well spread chromosomes were captured at a magnification of x 100 using the oil immersion objective to produce greater depths of focus. Chromosome counts were taken from 310 good metaphase cells. Measurements such as long and short arm lengths were made for each cell. Subsequently, calculations were made to determine the total chromosome length, arm ratio (s/l), ratio of short arm to the total chromosome length expressed as a percentage (F%), and ratio of sum of short arm length to the sum of total chromosome length relation in percentage (TF%). The karyotypes were constructed into metacentric, submetacentric and subtelocentric chromosomes using data obtained from measurements (Ojeba and Torres, 1996). RESULTS The characteristics of the four clones of G. latifolia used for the experiment are represented in Table 1. The clones were selected on the basis of flower possession and colour. The distinct flower colours were yellowish- purple, yellow, and yellowish-green for IMS-20-NJIABA, AKS-33-EKPENE EDIENE and ANS-38-AWKA, respectively (Plate 1). ENS-47-MBU does not flower. Other growth parameters of the different clones showed that clone ENS-47-MBU that does not flower had the highest leaf size, while ANS-38-AWKA had the least. Clones IMS-20-NJIABA and AKS-33-EKPENE EDIENE have very close values of inflorescence length and number of florets per inflorescence as well as equal number of matured follicles per inflorescence and viable seeds per follicle. Photomicrographs of the four clones of the plant species are presented in Plate 2. A1 and A2 represents IMS-20-NJIABA; B1 and B2, AKS-33-EKPENE EDIENE; C1 and C2, ANS-38-AWKA, D1 and D2, ENS-47-MBU metaphase chromosomes, respectively. The photomicrographs showed a somatic chromosome number of 2n = 16, 2n =16, 2n = 24, and 2n = 20 in IMS-20-NJIABA, AKS-33-EKPENE EDIENE, ANS-38-AWKA and ENS-47-MBU, respectively. Details of the karyotype with respect to chromosome lengths, lengths of long and short arms, arm ratio, F%, and TF% are presented in Tables 2, 3, 4, 5. In IMS-20-NJIABA, chromosome lengths ranged from 1.91-5.78 µm with a mean length of 4.11µm and standard deviation (SD) of 1.19. AKS-33-EKPENE EDIENE had chromosomes ranging from 2.40-6.22 µm with a mean length of 4.28µm and SD of 1.37. ANS-38-AWKA chromosomes ranged from 3.46-5.01µm, while those of ENS-47-MBU varied from 2.79-5.75 µm. The arm ratios varied from 0.43-0.96, 0.51-0.95, 0.28-0.95, and 0.51-0.90 in IMS-20-NJIABA, AKS-33-EKPENE EDIENE, ANS-38-AWKA and ENS-47-MBU, respectively. The F% values ranged from 36.97-49.82 in IMS-20-NJIABA, 33.85-48.77 in AKS-33-EKPENE EDIENE, 21.76-49.87 in ANS-38-AWKA, and 33.57-47.46 in ENS-47-MBU, respectively (Tables 2, 3, 4, 5). The somatic chromosome complement of IMS-20-NJIABA were composed of 6 metacentric and 2 submetacentric pairs; AKS-33-EKPENE EDIENE had 5 metacentric and 3 submetacentric pairs; ANS-38-AWKA had 5 metacentric, 2 submetacentric, and 1 subtelocentric pairs; whereas, ENS-47-MBU comprised 7 metacentric and 3 submetacentric pairs. Table 6 shows the karyotype formula and total chromosome length of the four clones of G. latifolia. With respect to total cellular chromatin length, the maximum (48.38µm) and minimum (32.85µm) values were recorded in ANS-38-AWKA (triploid) and IMS-20-NJIABA (diploid), respectively. AKS-33-EKPENE EDIENE and ENS-47-MBU fell within this range. DISCUSSION The photomicrographs showed that 2n = 16 is the diploid number of G .latifolia. This was observed in IMS-20-NJIABA and AKS-33-EKPENE EDIENE. The results collaborates with earlier findings of Ying et al. (1977) who reported that the family Asclepiadaceae had chromosome numbers ranging from x = 8 to 11 or 12. ANS-38-AWKA and ENS-47-MBU are polyploids of different categories. ANS-38-AWKA manifested as a triploid (2n = 3x =24) while ENS-47-MBU is a double-tetrasomy (2n = 2x + 2 + 2 = 20). The occurrence of these polyploids may be attributed to failure of normal mitotic division in a diploid body cell leading to increased sets of chromosomes (Uguru, 2004) or may be due to the presence of more than one nucleus per cell (Partanen, 1963). Polyploidy in some instances is advantageous as it affects plant part sizes like larger leaf areas, flowers and fruits (Walker et al. 2005; El-Ferchichi et al. 2006; Samiha et al. 2009). The higher number of viable seeds per follicle and matured follicles per inflorescence in IMS-20-NJIABA and AKS-33-EKPENE EDIENE may be attributed to the highly stable nature of diploid species in contrast to polyploids that are often plagued with instability and sterility (Uguru, 2004). ANS-38-AWKA (triploid) showed high level of sterility by having 50% less of viable seeds compared to IMS-20-NJIABA and AKS-33-EKPENE EDIENE. On the other hand, ENS-47-MBU does not flower however; it had the highest leaf area index. The sizes of chromosomes varied slightly with respect to the position of the centromere and arm lengths. Such variations among co-species may arise because of differences in the repetitive DNA sequences, which eventually make up the genome size (Schmidt et al., 1998). Karyotypic evidence revealed the preponderance of metacentric chromosomes with both arms approximately equal in all the clones. Others were sub-metacentric having one arm approximately twice the other with an exception to ANS-38-AWKA that had a pair of subtelocentric chromosome. This symmetry in morphology is a reflection of the relatively primitive karyotypes of the members of the species (Obute et al., 2005), an indication that species of G. latifolia is still going through the process of evolution. Even so, Davis and Heywood (1973) asserted that this view might not be universal since many highly evolved species are also known to show symmetry. Karyotype variation is important not only as a genetic variability but also because they represent an important micro-morphological feature for this species, and are frequently associated with species differentiation (Stebbins, 1971). The clones were characterized by a graded karyotype into medium small (B-type chromosome with average length of 2-5 µm) sized chromosomes. This is in accordance with the classification criteria proposed by Stebbins (1938). Furthermore, Candan and Kesereioglu (2009) reported that adaptation of species to environmental conditions is good when B-chromosomes are less whereas sterility increases in species with high B-chromosomes despite good environmental adaptation. Earlier, Rick (1987) reported significant correlation of leaf striping and or narrowing of leaves to increased number of B-chromosomes in maize. The results of low setting of florets after flowering in the species depicts low fertility as only two to four follicles survived to maturity in the three flowering species out of a range of 190-290 florets on an inflorescence in the three clones. This low fertility status of the species agrees with the reports of Candan et al. (2009) and Rick (1987) on the influence of B-chromosomes on fertility rate in plant species. The chromosomes of the polyploids were comparatively smaller in size than their diploid counterparts. This has been reported for polyploids in other dicotyledons (Stebbins, 1938). ENS-47-MBU had the highest total haploid chromatin length (40.9µm), while ANS-38-AWKA (triploid) had the least total haploid chromatin length (32.3µm). The morphological characters of the studied clones indicated that the polyploidy observed in ENS-47-MBU resulted in higher leaf size and its inability to flower. Furthermore, the triploid nature of ANS-38-AWKA resulted in reduction in all the measured metric traits and fertility. The result agrees with reports of other researchers in other plants (Stebbins, 1949; Allard, 1980). The chromosome sizes ranged from 1.91-6.22µm which is small compared to 14.4-85.5µm reported for Vigna unguiculata (Barone and Saccardo, 1990). The F% values implied slight variations in karyotypes. The minimum range (36.17 - 49.01%) was recorded in IMS-20-NJIABA, while the maximum range (21.76 - 49.87%) was recorded in ANS-38-AWKA due to the presence of a pair of subtelocentric chromosome. AKS-33-EKPENE EDIENE and ENS-47-MBU were in- between, having F% values of 33.85 - 48.77% and 33.57 - 47.46%, respectively. Therefore, on the basis of F% values and position of the primary constriction, IMS-20-NJIABA, AKS-33-EKPENE EDIENE, and ENS-47-MBU appear to manifest more primitive conditions than ANS-38-AWKA, which had a wider range of F%. This agrees with Stebbins (1971) who reported that species with more symmetry are primitive. The TF% value was between 40.99 - 45.29%, F% and TF% are two important parameters used in determining the position of the genotypes in evolution (Sinha and Roy, 1979). These results tend to suggest that the genotypes of G. latifolia are still going through the process of evolution. CONCLUSION This work has shown that the diploid number of chromosomes in G. latifolia is 2n = 16. The existence of polyploidy of different categories in some of the clones of G. latifolia is an indication that the chromosomes of G. latifolia is amenable to manipulations, and this might pose problems to chromosome pairing during hybridisation. However, the discovery of polyploidy, especially in ENS-47-MBU with the highest leaf area index is advantageous to crop breeders and consumers because of the possible exploitation of the broader leaf sizes associated with the clone. The non-flowering nature of the clone is not a disadvantage per se, as the plant can be mass propagated by vegetative cuttings. The metacentric and sub-metacentric morphology that predominates the clones indicates that species of G. latifolia has not yet fully evolved. However, the variations and B-size chromosomes observed in karyotypes of the plant species coupled with their different genomic levels reduces the chances of achieving high rates of successful crosses within the species as successful crosses will only be possible with plants of the same ploidy. Selection of clones with desired traits of interest from large populations will enhance faster improvements in the species especially as the plant is propagated vegetatively. REFERENCES
Copyright 2011 - African Crop Science Journal The following images related to this document are available:Photo images[cs11003t3.jpg] [cs11003t1.jpg] [cs11003p2.jpg] [cs11003t5.jpg] [cs11003f1.jpg] [cs11003t4.jpg] [cs11003p1.jpg] [cs11003t6.jpg] [cs11003t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}