 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 3, pp. 137-147 Agrobacterium-mediated transformation of common bean N. O. Amugune, B. Anyango and T.K. Mukiama School of Biological Sciences, University of Nairobi, P. O. Box 30197, Nairobi, Kenya
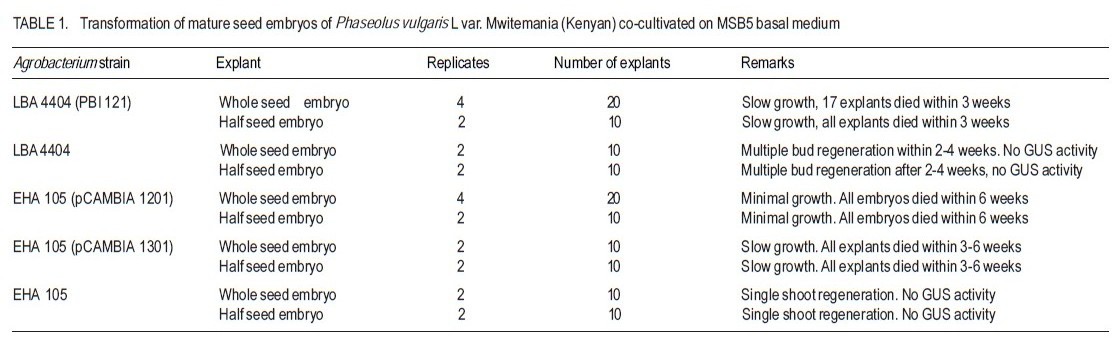
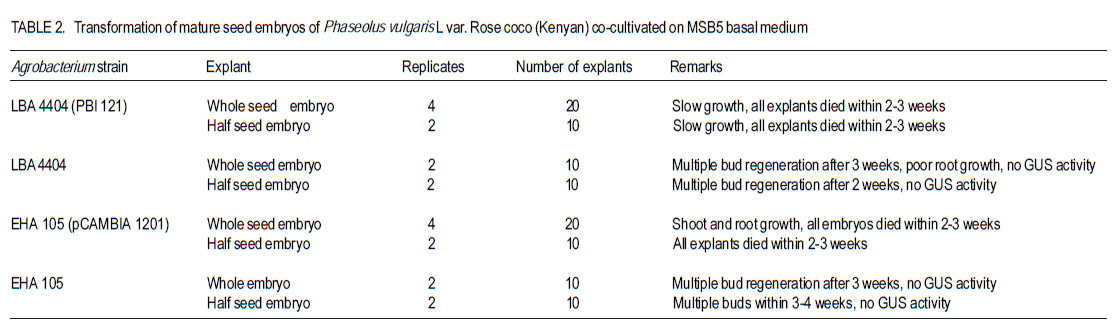
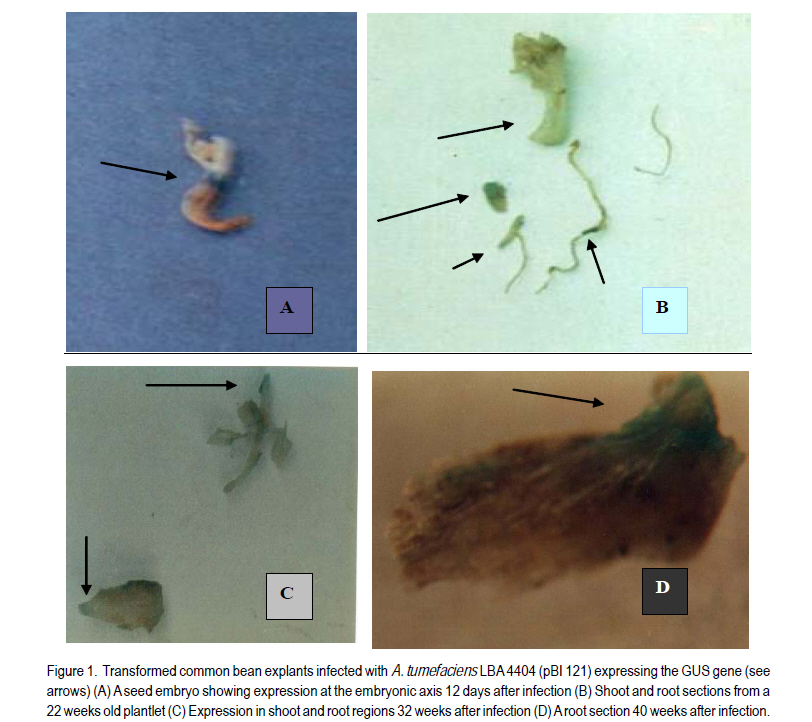
Received 4 April, 2011; accepted 12 September, 2011 Code Number: cs11014 ABSTRACTThe common bean (Phaseolus vulgaris L.) is an important human dietary constituent being a rich source of protein. Genetically improved bean varieties are required as optimum yields are not realised due to constraints such as diseases and insect pests. The objective of this study was to evaluate the potential of two common bean varieties Mwitemania and Rose coco to in vitro Agrobacterium tumefaciens- mediated transformation. Mature seed embryos germinated for 1-2 days on moist filter paper, were stab inoculated with A. tumefaciens strains LBA 4404 (pBI 121), EHA 105 (pCAMBIA 1201) and EHA 105 (pCAMBIA 1301), harbouring β-glucuronidase (GUS) intron plasmids. The infected embryos were co-cultivated for 3-4 days on basal Murashige and Skoog, 1962 medium with B5 vitamins (MSB5) or medium supplemented with 10 µM benzyl-aminopurine (BAP) and cultured on regeneration and selection medium consisting of 10 µM BAP and 50 mg L-1 kanamycin or hygromycin. Transformed shoots and roots confirmed by histochemical staining for GUS activity were obtained in 40 weeks old Mwitemania plantlets from explants infected with A. tumefaciens LBA 4404 (pBI 121). No GUS expression was observed in all Rose coco and Mwitemania shoots from explants infected with EHA 105 (pCAMBIA 1201) or EHA 105 (pCAMBIA 1301). Key Words: Agrobacterium tumefaciens, ß-glucuronidase, Phaseolus vulgaris RÉSUMÉ Le haricot commun (Phaseolus vulgaris L.) est un important constituant du regime alimentaire humain riche en protéines. Des variétés génétiquement améliorées sont nécessaires, les rendements optimum n’étant pas realisés suite aux contraintes comme maladies et pestes dues aux insects. L’objectif de cette étude était d’évaluer le potential de deux variétés de haricot commun, Mwitemania et Rose coco en transformation médiée in vitro Agrobacterium tumefaciens. De grains embryonnaires matures en germination sous papier filter pendant 1-2 jours, étaient inoculés en stab avec des strains de LBA 4404 (pBI 121) A. tumefaciens, EHA 105 (pCAMBIA 1201) et EHA 105 (pCAMBIA 1301), harbouring β-glucuronidase (GUS) “intron plasmids”. Lews embryons infectés étaient c0-cultivés pendant 3-4 jours sur la base de media Murashige and Skoog, 1962 avec des vitamins B5 (MSB5) ou media supplémenté avec 10mM benzyl-aminopurine (BAP) et cultivé sur le média sélectionné et régénéré composé de 10 mM BAP et 50 mg L-1 kanamycin ou hygromycin. Les tiges et racines transformées, confirmées par la culture histochimique pour activité GUS étaient obtenues dans les plantules de Mwitemania âgées de 40 semaines issues des explants infectés avec A. tumefaciens LBA 4404 (pBI 121). Aucune expression GUS n’était observée dans toutes les tiges de Rose coco et Mwitemania shoots issues des explants infectés avec EHA 105 (pCAMBIA 1201) ou EHA 105 (pCAMBIA 1301). Mots Clés: Agrobacterium tumefaciens, ß-glucuronidase, Phaseolus vulgaris INTRODUCTION The common bean (Phaseolus vulgaris L.) is the most important food legume worldwide and a leading source of low cost proteins (Arellano et al., 2009). Despite having a potential yield of over 4000 kg ha-1, bean yields in West Asia and North America range from 1100 to 1500 kg ha-1 while yield in Latin America and African countries range from 500 to 600 kg ha-1 (Aragao and Rech, 2001). Some of the factors contributing to low yields are poor agronomic practices, diseases and insect pests, low soil fertility and lack of improved varieties (Aragao et al., 2002). Besides the conventional breeding techniques, attempts have been made to obtain improved bean varieties through genetic transformation. Two methods are currently available for common bean transformation, indirect gene transfer using a disarmed gram negative soil bacterium such as Agrobacterium, or direct gene transfer techniques mainly particle bombardment or electroporation (Dillen et al., 1995; Kim and Minamikawa, 1997; Aragao et al., 2002; Rech et al., 2008). Transformation can be monitored by use of the β-glucuronidase (GUS) gene of Escherichia coli (Jefferson, 1987). The GUS, intron gene allows expression in transformed plant tissues but not in bacterial cells and can be used for direct determination of the transformation rate as a measure of susceptibility. Histochemical GUS assays (Jefferson, 1987) and green fluorescent protein (GFP) techniques (Zambre et al., 2005; Colpaert et al., 2008) are available to detect specific cell type gene expression. The common bean, like other legumes, is generally recalcitrant to Agrobacterium-mediated transformation due to poor regeneration in tissue culture (Svetleval et al., 2003; Zambre et al., 2005; Colpaert et al., 2008; Arellano et al., 2009). Most grain legumes transformed by Agrobacteriumbased methods generally yield low rates of transformation, for instance, 0.1- 5 % (Atkins and Smith, 1997; Veltcheva et al., 2005). Some leguminous species that have been transformed using Agrobacterium include soybean, Glycine max. Mer. (Chee et al., 1989; Liu et al., 2004; Paz et al., 2005; Dang and Wei, 2007), pea nut, Arachis hypogea L. (Eapen and George, 1994), pea, Pisum sativum (Puonti- Kaelas et al., 1990; Schroeder et al., 1993) and cow pea, Vigna unguiculata L (Polpelka et al., 2006). Bean explants that have been used in Agrobacterium-mediated transformation include cotyledonary nodes (McClean et al., 1991), protoplasts (Leon et al., 1991), mature seed embryos (Aragao et al., 1992), leaf discs and hypocotyls (Franklin et al.,1993), seed meristems (Russell et al., 1993), intact shoot tips of germinating seeds (Lewis and Bliss, 1994), shoot apex of embryogenic axis (Kim and Minamikawa, 1995), apical meristems (Brasileiro et al., 1996), seedlings and shoot apexes (Aragao and Rech, 1997), and embryo axis (Dillen et al., 1995; 1997). Kanamycin was used in some cases as the selective agent and transformation was confirmed using GUS expression (Aragao et al., 1993). McClean et al. (1991) obtained transformed callus and roots from cotyledonary nodes and hypocotyls of dry bean infected with A. tumefaciens strain C58Z707 and avirulent A. rhizogenes strain A4RS (pRiB278b), respectively. The transformed callus and roots, though capable of growing on MS medium with 5 µM BAP and 50 mg L-1 kanamycin could not regenerate plants. Franklin et al. (1993) produced a kanamycin resistant callus from leaf disc or hypocotyl explants of green bean infected with A. tumefaciens EHA 101 (pKYLX71GUS) using SH (Schenk and Hildebrant, 1972) media with 10 µM p-chlorophenoxy acetic acid (pCA), 2 µM 2,4 – dichlorophenoxyacetic acid (2,4 –D) and 0.5 µM kinetin (KIN). The callus expressed βglucuronidase (GUS) activity but no transgenic plants were obtained. Dillen et al. (1997) obtained GUS expression in regenerated bean shoots derived from seedling node explants infected with A. tumefaciens - A2760 (Hoekema et al., 1983) and EHA 105 (Hood et al., 1993) using an introncontaining GUS gene as a reporter system. However, no transgenic common bean plants were recovered. Zambre et al. (1998; 2005) using Agrobacterium obtained transgenic plants in Adzuki bean, Phaseolus acutifolius but not in P. vulgaris. The axillary bud (Franklin et al., 1993; Zhang et al., 1997) and embryonic axis (Gatica-Arias et al., 2010) tissue culture protocols for whole plant regeneration described in the common bean are yet to be adopted successfully for use in an Agrobacterium based transformation system, hence the need to explore other methods. Lewis and Bliss (1994) obtained tumours and GUS expression when intact shoot tips of germinating P. vulgaris seeds were stab inoculated in planta with A. tumefaciens C58, but no transgenic plants were recovered. Such in planta techniques overcome the complications of regenerating plants from tissue-cultured material, as regeneration does not rely on shoot production from callus or from suspensions of cells or protoplasts. In soybean (Glycine max) Chee et al. (1989) produced transgenic plants, and seedderived progeny in planta by infecting plumules, cotyledonary nodes and surrounding regions of axenic soy bean seedlings with A. tumefaciens carrying binary vectors. Colpaert et al. (2008) obtained transgenic roots expressing GUS and GFP fluorescence from de-rooted P. vulgaris seedlings infected in vitro with Agrobacterium rhizogenes. Stable transformation has been realised in common bean using the direct gene delivery systems. Transgenic common bean plants have been obtained by particle bombardment of DNA into mature seed meristems (Russell et al., 1993; Aragao et al., 1996). Transgenic bean plants expressing resistance to bean golden mosaic geminivirus (Aragao et al., 1998) and tolerance to the herbicides glufosinate ammonium and imazapyr (Aragao et al., 2002; Rech et al., 2008) have been developed through particle bombardment. The development of an efficient gene transfer system in the bean using A. tumefaciens would greatly benefit breeding programmes by introducing novel genes into the crop. The objective of the study was to determine whether mature zygotic embryos of two common bean varieties grown in Kenya could be transformed in vitro by Agrobacterium tumefaciens. MATERIALS AND METHODS Plant material and explant preparations. Mature seeds of two popular common bean varieties grown in Kenya, Rose coco (GLP-2) and Mwitemania (GLP-92), were surface sterilised in 70% (v/v) ethanol for 2 minutes and 1 % (v/v) sodium hypochlorite for 20 minutes. The sterilised seeds were rinsed four times with sterile water and germinated on moist sterile filter paper for 1 to 2 days or until the radicle emerged. The cotyledons of the germinating seeds were split open to expose the embryo, which was then detached. Half seed embryos were obtained by the removal of the root apex as described by Schroeder et al. (1993). Medium. Murashige and Skoog (1962), medium supplemented with B5 vitamin (Gamborg et al., 1968) was used in all the experiments. The medium abbreviated as MSB5, consisted of MS macroand micro-salts plus 30 g L-1 sucrose, 10 mg L-1 thiamine HCl, 1 mg L-1 pyridoxine HCl; 1 mg L-1 nicotinic acid and 100 mg L-1 myo-inositol. The pH was adjusted to 5.8± 0.1 using 1.0 M hydrochloric acid or sodium hydroxide and the medium solidified by adding 8 g L-1 agar. The medium was sterilised by autoclaving at 1 kg cm-2 and 121 °C for 20 minutes. Thereafter, the medium was dispensed into sterile bottles or petri dishes in a laminar flow cabinet. Preparation of A. tumefaciens cultures. Solid and suspension cultures of Agrobacterium tumefaciens strains: LBA 4404 (pBI 121), EHA 105 (pCAMBIA 1201) and EHA 105 (pCAMBIA 1301), harbouring β-glucuronidase (GUS) intron plasmids were used. A colony of A. tumefaciens LBA 4404 (pBI 121) was streaked onto solidified Luria Bertani (LB) medium (Miller, 1972) containing 100 µgml-1 rifampicin and 100 µgml-1 kanamycin and incubated at 28 °C for 3 days. Liquid A. tumefaciens LBA 4404 (pBI 121) cultures were initiated by picking off a colony with a needle, and inoculating 125 ml LB medium containing 100 µgml-1 rifampicin and 100 µg ml-1 kanamycin in 250 ml conical flasks. Cell suspensions were grown overnight on a shaker at 28 °C and 180 revolutions per minute. A colony of A. tumefaciens, EHA 105 (pCAMBIA 1201), was streaked onto solidified LB medium containing 100 µgml-1 chloramphenicol and incubated at 28 °C for 2 - 3 days. Thereafter, liquid cultures were prepared by inoculating non-solidified LB medium containing 100 µgml-1 chloramphenicol with a colony of A. tumefaciens EHA 105 (pCAMBIA 1201) and incubating overnight on a shaker as already described. A. tumefaciens EHA 105 (pCAMBIA 1301) solid and liquid cultures were prepared in a similar manner except that 50 µgml-1 kanamycin was used for bacterial selection instead of 100 µgml-1 chloramphenicol. The negative control A. tumefaciens LBA 4404 and EHA 105 were grown on LB media without antibiotics. In vitro infection of seed embryos. Whole mature germinated embryos of the two bean genotypes were infected by injecting the embryonic axis using a 30-gauge needle attached to a 1-ml syringe containing liquid Agrobacterium culture as described by Lewis and Bliss (1994). Half seed embryos with the root apices removed, were inoculated by dipping in each bacterial suspension culture for 5 to 15 minutes and then dried using sterile tissue paper to remove excess bacteria. The infected explants were co-cultivated on MSB5 basal or medium supplemented with 10 µM BAP for three to four days. After cocultivation, explants infected with A. tumefaciens LBA 4404 (pBI 121) were transferred onto MSB 5 medium supplemented with 10 µM BAP, 50 mg L-1 kanamycin and 250 mg L-1 cefotaxime for shoot growth and selection of transformed plants. Cefotaxime was used to kill excess bacteria. Those infected with A. tumefaciens EHA 105 (pCAMBIA 1201) and EHA 105 (pCAMBIA 1301) were transferred onto medium supplemented with 10 µM BAP, 50 mg L-1 hygromycin and 250 mg L-1 cefotaxime. Co-cultivated control explants infected with A. tumefaciens LBA 4404 and EHA 105 were transferred onto medium with 10 µM BAP and 250 mg L-1 cefotaxime. All the cultures were incubated at 25 ± 2 º C, under a 16 / 8 hour dark light cycle and 3000 lux light intensity, respectively. β - Glucuronidase (GUS) assay. Explants of the two bean genotypes at various stages of growth were tested for β-glucuronidase (GUS) activity using the histochemical staining method described by Jefferson et al. (1987). The first histochemical assay was done after 12 days using three explants removed at random from each replicate. The GUS assay was repeated after 22, 32 and 40 weeks. RESULTS Mature whole seed embryo explants of P. vulgaris L var. Mwitemania infected with strain LBA4404 (pBI 121) and co-cultivated on MSB5 basal medium exhibited slow growth when subcultured onto MSB5 medium supplemented with 10 µM BAP, 50 mg L-1 kanamycin and 250 mg L-1 cefotaxime(Table 1). Out of the 20 whole mature seed embryos infected, 17 explants (85%) died within three weeks and the remaining three (15%) died after 10 weeks. All the 10 half seed embryos also infected with strain LBA 4404 (pBI 121) died within three weeks. The control, whole and half mature seed embryo explants infected with strain LBA 4404 and cultured on medium with 10 µM BAP and 250 mg L-1 cefotaxime, regenerated multiple buds after two to four weeks but these were confirmed to be negative for GUS activity when tested after three to 10 weeks. Mature whole and half seed embryos of var. Mwitemania co-cultivated on MSB5 basal medium after infection with either A. tumefaciens strain EHA105 (pCAMBIA) 1201 or EHA 105 (pCAMBIA 1301) died within six weeks of culture on to MSB5 medium with 10 µM BAP, 250 mg L-1 cefotaxime and 50 mg L-1 hygromycin (Table 1). The control explants infected with strain EHA 105 did not regenerate multiple buds, but gave rise to single shoot and root meristems, which were negative for GUS activity when tested after eight to ten weeks. Table 2 shows data for whole and half mature seed embryos of P. vulgaris var. Rose coco, cocultivated on MSB5 basal medium for three days after infection with A. tumefaciens LBA 4404 (pBI 121) and EHA 105 (pCAMBIA 1201). All the explants died within two to three weeks when cultured on MSB5 selection medium supplemented with 10 µM BAP, 50 mg L-1 kanamycin or hygromycin and 250 mg L-1 cefotaxime. Thirteen out of 20 whole mature seed embryos of var. Rose coco co-cultivated on MSB5 basal medium after infection with strain EHA 105 (pCAMBIA 1201) turned green within 5 days, but exhibited minimal shoot growth during the first 2 weeks of culture onto MSB5 medium supplemented with 10 µM BAP, 50 mg L-1 hygromycin and 250 mg L-1 cefotaxime. Root regeneration was observed in 3 out 20 explants within the same period. The half seed embryos infected with the same strain and were cultured on the same medium failed to show any significant growth, and died within 3 weeks. The control whole and half seed embryos infected with A. tumefaciens strains LBA 4404 and EHA 105 regenerated multiple buds within 2-4 weeks (Table 2). In variety Mwitemania 33 mature whole seed embryos infected by stab inoculation with A. tumefaciens LBA 4404 (pBI 121) and co-cultivated on MSB5 medium supplemented with 10 µM BAP for three days turned green after nine days of culture on medium with 10 µM BAP, 50 mg L-1 kanamycin and 250 mg L-1 cefotaxime (Table 3). The explants exhibited single shoot meristem growth in addition to root regeneration. One out of three explants selected at random after 12 days of culture on shoot regeneration and selection medium exhibited GUS activity evidenced by the blue staining at the embryonic axis (Fig. 1 A). Fourteen out of the 30 remaining embryo explants died within 20 weeks. Eleven out of the 16 surviving embryo explants exhibited shoot growth, three showed both shoot and root growth while two failed to show any significant growth of either the shoot or root region despite regular subculture to shoot regeneration and selection medium of the same composition every two to three weeks. The number of surviving embryo explants from those initially infected (Table 3) continued to decline in subsequent subcultures and only nine regenerated to form plantlets on shoot induction and selection medium consisting of 10 µM BAP, 50 mg L-1 kanamycin and 250 mg L-1 cefotaxime after 22 weeks. Two out of the nine plantlets removed at random were both positive for GUS activity when tested after 22 weeks. However, the transformation was chimaeric as both the shoot and the root regions showed partial expression of the GUS gene (Fig. 1 B). Five out of the seven 30 weeks’ old plantlets subcultured onto a fresh shoot induction and selection medium exhibited normal shoot growth but root growth was poor. There was also presence of GUS activity in both the shoot and root segments of two plantlets selected at random when tested after 32 and 40 weeks (Fig. 1 C and 1 D). Two out of 28 (7.1%) embryo explants from the lot initially infected with A. tumefaciens LBA 4404 (pBI 121) excluding those used in the GUS assay exhibited growth on shoot regeneration and selection medium after 40 weeks. However, the two plantlets eventually died when transferred to vermiculite for hardening; therefore, mature plants were not raised. The negative control explants infected with A. tumefaciens strain LBA 4404 and were cultured on shoot regeneration medium consisting of 10 µM BAP and 250 mg L-1 cefotaxime after co-cultivation on medium with 10 µM BAP exhibited shoot and root meristem growth and developed multiple buds after 12 days. There was no GUS activity when the explants were tested after nine days, as well as after 21 and 32 weeks. All the 28 whole mature seed embryos explants of var. Mwitemania, infected with A. tumefaciens strain EHA 105 (pCAMBIA 1301) and were cultured on MSB5 medium with 10 µM BAP, 50 mg L-1 hygromycin and 250 mg L-1 cefotaxime after co-cultivation on medium with 10 mM BAP, died within 4 weeks (Table 3). However, the 24 negative control explants infected with A. tumefaciens EHA 105 developed multiple buds within 12 days. There was no GUS activity in the shoots regenerated from negative control explants when tested after two and six weeks. DISCUSSION Mature seed embryos of variety Mwitemania (Table 1) and Rose coco (Table 2) that were stab inoculated with A. tumefaciens LBA 4404 (pBI 121), but were co-cultivated on MSB5 basal medium could not be transformed. The absence of growth hormones during co-cultivation of explants and poor regeneration in P. vulgaris resulted in very limited growth, which may have been insufficient for transformation to occur. A suitable regeneration system is necessary for recovery of transformed plants as Agrobacterium usually damages plant tissues during infection (Zhang et al., 1997). Zygotic bean embryo explants infected with either A. tumefaciens EHA 105 (pCAMBIA 1201) or A. tumefaciens EHA 105 (pCAMBIA 1301) exhibited limited growth and died within six weeks. This was probably because the 50 mg L-1 hygromycin used for selection may have been toxic to the tissues subsequently making regeneration of multiple buds difficult. Also strain EHA 105 though previously used in rice transformation (Hiei et al., 1994) may be unsuitable for common bean transformation. Evidence of transformation of bean varieties was shown by the fact that infected seed embryo explants co-cultivated on medium with 10 µM BAP could be grown on shoot regeneration and selection medium consisting of 10 µM BAP, 50 mg L-1 kanamycin and 250 mg L-1 cefotaxime for 40 weeks (Table 3). The transformed seed embryos developed directly into a single shoot and root system and exhibited GUS activity. The expression of GUS activity in the shoot region of germinating P. vulgaris var.Mwitemania seed embryo explants infected with A. tumefaciens LBA 4404 (pBI 121) was an indication that transgenic plants can be recovered. The noncontinuous position of the blue staining zones in the bean shoot indicated that the transformation was chimaeric as none of the events was in continuing cell lineages of the meristem. The wound site created by stabbing the zygotic bean explants was necessary for Agrobacterium infection. It also exposed the interior layers of the meristem to Agrobacteria infection. Transformation of these layers would allow a subsequently arising germline to pass on the acquired transgene through the seed (Lewis and Bliss, 1994). Wounded plant tissues also produce phenolic compounds necessary for Agrobacterium infection (Kado, 1991). Presence of the growth regulator BAP in both co-cultivation and shoot regeneration media greatly enhanced transformation and the survival rate of the transformed bean explants. This result is in agreement with previous studies in which presence of growth regulators in co-cultivation medium has been shown to enhance transformation in the common bean (Zhang et al., 1997), and in pea, Pisum sativum (Schroeder et al., 1993). In previous studies, stab inoculation of intact meristems led to recovery of transgenic soyabean (Chee et al., 1989), but not common bean. Zhang et al. (1997) demonstrated that in P. vulgaris the explant tissue, the Agrobacterium strain and the presence of growth regulators in the co-cultivation medium were found to be important in transformation. Transformation of common bean with the particle gun though has yielded transformed plants (Aragao et al., 1996; 2002; Rech et al., 2008), may produce lower levels of stable inheritance and is more expensive when compared to the Agrobacterium method. Despite the fact that transgenic bean plants were not recovered, the stab inoculation method described is a promising one and requires further study as there are limited reports on successful transformation of P. vulgaris using Agrobacterium. Also, reducing the period of bean explant exposure to selection medium containing antibiotics to three to four weeks could lead to recovery of transformed plants as in the case of biolistic transformation (Aragao et al., 2002; Rech et al., 2008). CONCLUSION Mature seed embryos of Phaseolus vulgaris var. Mwitemania can be transformed by Agrobacterium tumefaciens. Of the vectors tested, A. tumefaciens LBA 4404 (pBI 121) transforms the bean embryos. Histochemical staining for GUS activity has confirmed expression in the germinating seed embryos of variety Mwitemania infected with A. tumefaciens LBA 4404 (pBI 121). In order for transformation to occur, cytokinin is necessary in both the media used for co-cultivation and selection of transformed plants. ACKNOWLEDGEMENT We thank Dr. D. Burger, of ARC Roodeplaat Vegetable and Ornamental Plant Institute, Republic of South Africa for supplying the Agrobacterium tumefaciens strains LBA 4404 and LBA 4404 (pBI 121). Dr. R. Harcout, of the Centre for the Application of Molecular Biology to International Agriculture (CAMBIA), Australia supplied A. tumefaciens strains EHA 105, EHA 105 (pCAMBIA 1201) and EHA 105 (pCAMBIA 1301) used in the study. The Germany Government through DAAD provided financial support. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11014t3.jpg] [cs11014t1.jpg] [cs11014f1.jpg] [cs11014t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}