 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 3, pp. 149-163 Genetic analysis for high temperature tolerance in bread wheat S.S. Punia, A. Mansoor Shah and B. Ram Ranwha Department of Plant Breeding and Genetics, Rajasthan College of Agriculture, Maharana Paratap University of Agriculture &Technology, Udaipur-313001, Rajasthan, India Corresponding author: sumerspunia@hotmail.com Received 10 February, 2011; accepted 12 September, 2011 Code Number: cs11015 ABSTRACT Heat stress, characterised by a trend in average temperature increase during anthesis and grain filling, leads to forced maturity is one of the major constraints of wheat production in arid and, semiarid regions of the world. This study examined the nature and magnitude of gene action for yield and its contributing characters and some important heat tolerant parameters in bread wheat (Triticum aestivum l em. thell) to determine breeding strategies for future breeding programmes. Twelve lines and four testers were crossed in an L x T mating design. The 48 crosses and their parents were raised under normal (21, November 2002) and late sown (1, January 2003) seasons at the Experimental Farm of Rajasthan College of Agriculture, Udaipur in India. Lines K’sona, DWR 195, C 306 and K 9708 were found to be good combiners for different heat tolerant parameters along with grain yield. Combinations of Raj 3077 x Kailash under normal (E1) and late sown (E2), C 306 x PBN 51 (E1) and C 306 x HD 2189 (E2) for grain yield; had high sca effect. The crosses viz., K 9708 x PBN 51 for proline content and heat injury, DWR 195 with HD 2189, and C 306 with Kailash for heat injury had desirable significant sca effects. The other crosses, HD 2329 x Kailash for pollen viability, and Raj 3765 x Kailash for chlorophyll content, were the best specific combiners. Parents like C 306, K’sona, DWR 195, K 9708, Raj 3077, PBN 51 and Kailash could be utilised in multiple crossing programmes and further biparental matting for selection of high yielding progenies for heat tolerance. Key Words: Heat tolerant, proline, Triticum aestivum RÉSUMÉ Le stress dû à la chaleur, caractérisé par une tendance à l’augmentation de la température moyenne au cours de l’anthésis et du remplissage de grains, et conduisant à la maturation forcée est une des contraintes majeures à la production du blé dans des régions arides et semi arides du monde. Cette étude a examiné la nature et la magnitude de l’action de gènes de rendement et ses caractères contributifs ainsi que quelques paramètres importants de tolérance dans le blé patissier (Triticum aestivum) afin de déterminer des stratégies d’amélioration pour des programmes futures d’amélioration. Douze lignées et quatre testeurs étaient croisés en dispositf L x T. Les 48 croisements et leurs parents étaient plantés en saisons normale (21, November 2002) et tardive (1, January 2003) à la ferme expérimentale du collège d’Agriculture de Rajasthan, Udaipur en Inde. Les lignées K’sona, DWR 195, C 306 et K 9708 étaient jugées de bons combinants pour différents paramètres de tolérance à la chaleur avec un bon redement en grains. Les combinaisons de Raj 3077 x Kailash en saison normale (E1) et plantées tardivement (E2), C 306 x PBN 51 (E1) et C 306 x HD 2189 (E2) pour rendement en grain avaient un effet sca élevé.Les croisements telque K 9708 x PBN 51 avec for proline content and heat injury, DWR 195 avec HD 2189, etC 306 avec Kailash pour dommage de la chaleur avaient d’effects sca significatifs. Les autres croisements, HD 2329 x Kailash pour variabilité de pollen, et Raj 3765 x Kailash pour contenu chlorophyllien, étaient les meilleurs combinants spécifiques. Les parents dont C 306, K’sona, DWR 195, K 9708, Raj 3077, PBN 51 et Kailash pourraient être utilisés dans de multiples programmes de croisement et autres couvertures biparentales pour la sélection de progénies à rendement élevé avec tolérance à la chaleur. Mots Clés: Tolérance à la chaleur, proline, Triticum aestivum INTRODUCTION Wheat (Triticum aestivum l Em. Thell) is the most widely consumed cereal crop worldwide. Globally, demand for wheat by the year 2020 is forecast at around 950 million tonnes per year (Rosegrant et al., 1995; Kronstad, 1998). This target will be achieved only, if global wheat production is increased by 2.5% per annum. Wheat best adopts to cool growing conditions (Chowdhury and Wardlaw, 1978), while moderately high temperatures (25 -32 oC) for longer duration and very high temperature (33-40 oC) for a shorter period are very common in subtropical environments of South East Asia including India (Paulsen 1994; Stone and Nicholas, 1994). Although wheat production in much warmer areas is technically feasible, heat stress is a common constraint, especially during anthesis and grain filling in many temperate environments in South and West Asia (Reynolds et al., 1994).Therefore, heat stress is one of the major constraints of wheat production in arid, semiarid, tropical and subtropical regions of the world (Ashraf and Harris, 2005). Heat stress at late growth stages is a problem in 40% of wheat areas in the temperate environments (Reynolds et al., 2001). A brief period of exposure to high ambient temperature (>35 oC) can drastically reduce grain yield in wheat (Hawker and Jenner, 1993) because of induction of early senescence and acceleration of grain filling activities in wheat (Paulsen, 1994) due to shortening of grain filling duration and constriction of carbon assimilation (Stone, 2001). Grain weight is affected by high temperatures, especially those above 34 ºC, that reduce the duration of grain filling owing to the limited photosynthesis (Al Khatip and Paulsen, 1984), and inhibit starch biosynthesis in the endosperm (Keeling et al., 1993; Jenner, 1994). The optimum time for sowing wheat on the plains of South Asia is from November the 15 to 25th, but in a significantly large area, principally the fourteen-million hectare South Asian rice/ wheat cropping system (Pandey et al., 2005; Joshi et al., 2007a), wheat sowing often gets delayed due to the late harvesting of rice (Arun et al., 2003). In about four to five million hectares, wheat-sowing occurs after December 15th, resulting in high wheat yield losses due to the reduced growth and development period available before harvest (Arun et al., 2003; Joshi et al., 2007b). Almost the whole of this late sown area is in the eastern Gangetic plains, most of which occurs in the 9.5 million hectare North Eastern Plain Zone (NEPZ) of India, where the average wheat yield is only 2.7 tonnes ha-1 compared to the 4.7 t ha-1 in the Indian State of Punjab (Joshi et al., 2007b). The North Western Plains Zone of India (extending from the state of Punjab to western Uttar Pradesh) has an area almost equal to that of the NEPZ, and is the most important wheat-producing zone in Indian, with an average wheat yield of about 4 t ha-1. Heat stress during post-anthesis (grain-filling stage) affects availability and translocation of photosynthates to the developing kernels and starch synthesis and deposition within the kernel, thus resulting in lower grain weight and altered grain quality (Bhullar and Jenner, 1985; Mohammadi et al., 2004). It has been observed that each degree rise in ambient temperature reduces the yield by 3-4% (Mishra, 2007). Wheat breeders are seeking to incorporate late heat tolerance in the wheat germplasm and to develop genotypes that are early in maturity in order to escape the terminal heat stress and, thus suit well in the rice-wheat as well as in soybean-wheat cropping systems. Plant physiological processes differ in their response to heat stress from one phenological stage to another (Fischer, 1985). Breeders evaluate many lines during selection for heat tolerance because identification of a plant with all the required genes is difficult (Ortiz-Ferrara et al., 1993). Selection for yield with heat tolerance is difficult in large breeding programmes with thousands of segregating lines. Though, physiological and biochemical screening techniques, as a complement to empirical methods could increase selection efficiency, securing heat tolerance genes that may be lost during empirical selection (Reynolds et al., 1994). A number of high temperature stress-related traits have received considerable attention, in particular membrane thermostability (Saadalla et al., 1990), canopy temperature depression (Blum et al., 1982), proline content and chlorophyll content. Information on the genetic control of these traits would aid in choosing parents for heat tolerance breeding programmes. Therefore, this study was carried out to identify the best combining parents and their crosses and to know the genetic architecture and mode of inheritance of traits related to high temperature, yield and its contributing traits. MATERIALS AND METHODS Plant materials and experimental design. Sixteen genetically diverse varieties of bread wheat (Triticum aestivum L. em.Thell) comprising of twelve lines, Raj 3777, WH 542, HD 2329, Raj 3077, K 9708, C 306, Raj 1482, Kalyansona, GW 190, Raj 3765, DWR 195 and HI 977, and four testers HD 2189, Ajantha, PBN 51 and Kailash, were selected on the basis of heat and drought tolerance, earliness and ecogeographic origin. They were crossed in L x T mating design. The 48 hybrids produced and their parents (including checks) were raised in randomised complete block design with three replications, at the Experimental Farm of the Department of Plant Breeding and Genetics (24o 34' N, 37o 42' E, Rajasthan College of Agriculture, Udaipur, Rajasthan in India). This was done under two dates of sowing viz., normal (E1-21st November 2002) and late (E2- 1st January 2003). In each replication and sowing date, single rows of parents and F1s were sown in 3 m row length with spacing of 5 cm between plants and 22.5 cm between rows. Characters studied. Five competitive plants were chosen randomly from equally spaced plants from the parents and F1s in each replicate and environment for observations on plant height, number of productive tillers per plant, number of spikelets per spike, number of grains per spike, 100-grain weight and grain yield per plant. Days to 50% flowering and days to 75% physiological maturity were recorded on plot basis. Heat injury, stomatal frequency and chlorophyll content were recorded on normal environment; whereas proline content and pollen viability were recorded under both the environments (E1 and E2). For these parameters except pollen viability, three fully expanded flag leaves were taken randomly as samples at postanthesis in each treatment (parents and F1s). For the pollen viability test, pollen grains were collected randomly from three different plants for each treatments at the initiation of anthesis when nearly half portion of spike emerge from flag leaf. They were stained with potassium iodide (1 or 2% potassium iodide + iodine crystal) and observed under a microscope. The viable pollen grains were expressed as a percentage of pollen in the sample. Leaf canopy temperature depression was measured by Infrared Thermometry at grain filling stage. The heat injury was then calculated using Equation (1)
Where: T = Conductance values for treatment; Stomatal frequency (upper and lower) was calculated using Xylene Thermocol Print method, where a viscous base solution of xylene and thermocol was used. For preparation of this solution, about 100 mL-1 of xylene was taken in beaker and kept on hot a plate for gentle heating up to when the thermocol disolved. Then small pieces of thermocol were added in a beaker and stirred with a glass rod until a viscous liquid solution was obtained. After cooling this solution at room temperature, a thin layer of the base solution was applied in the middle portion of the leaf-lamina on upper as well as lower surfaces for the observation of stomatae. This leaf base solution structure was allowed to dry at room temperature (25 oC) for 3 to 5 minutes. The stomatae had printed on the thermocol coat, while xylene part of the base solution was absorbed by the leaf. After drying, the thermocol coat was peeled off from leaf lamina. The thermocol layer with stomatae imprints, were stained with acetocarmine dye and observed under a compound microscope under high magnificence. Chlorophyll content was estimated by the Dimethylformamide method (Morgan and Porath, 1980) using Equation 2.
The optical density reading of extract was recorded at 663 Ao nm and 645 Ao nm by spectrophotometer. The value 0.03 is the leaf sample size in grammes. Statistical analysis. Statistical analysis was done by using BR state software (Ranwha, 1995). The statistical model for analysis of variance for individual environment for the randomised block design followed was:
Where: Yij = value of ith genotype in the jth replication; µ = an effect, common to all genotypes over replication; Gi = an effect of ith genotype over the replication; Rj = an effect of jth replication and sij= an uncontrolled variation associated with ith genotype and jth replication. The combining ability analysis for line x tester mating design was performed as per the method suggested by Singh and Choudhary (1995).The effects of gca and sca were estimated as:
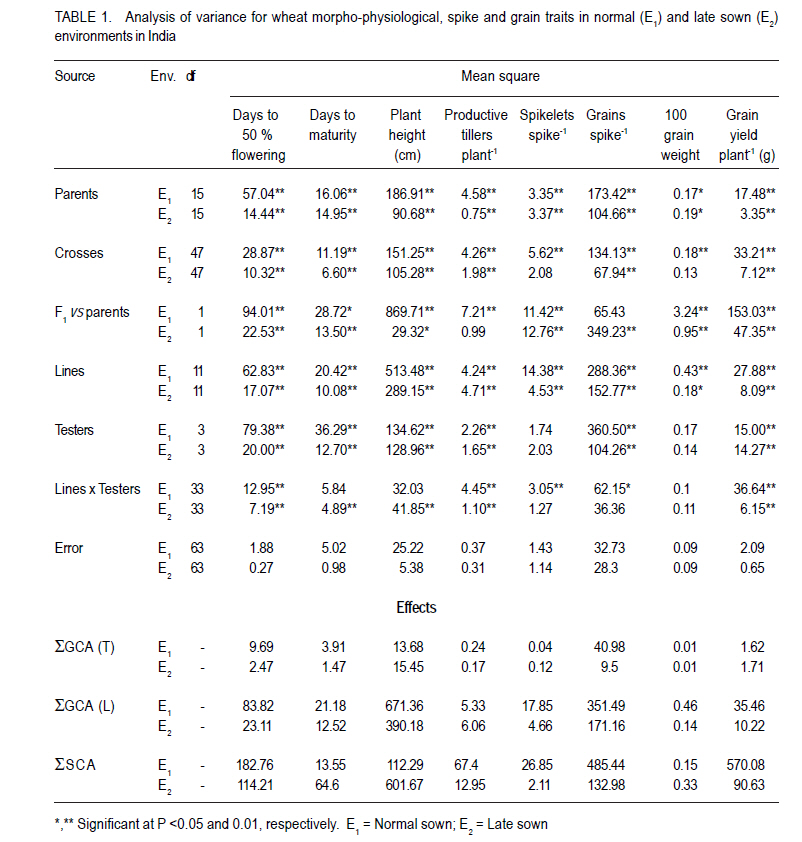
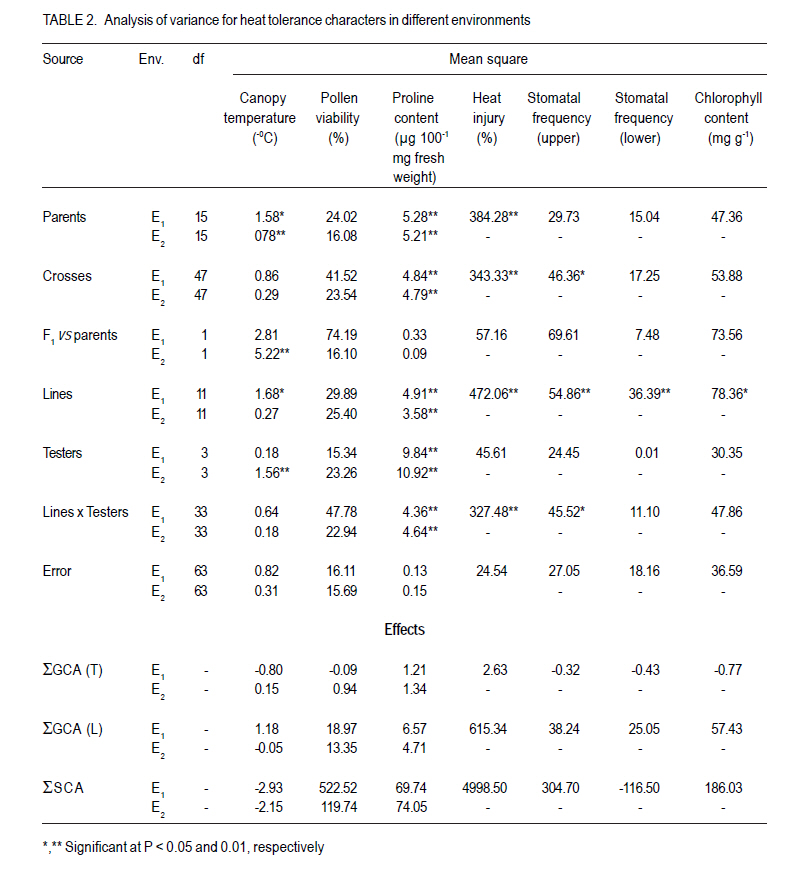
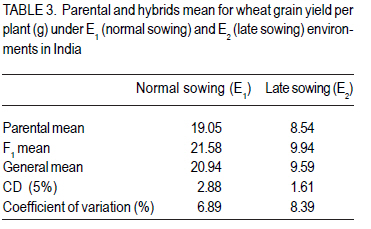
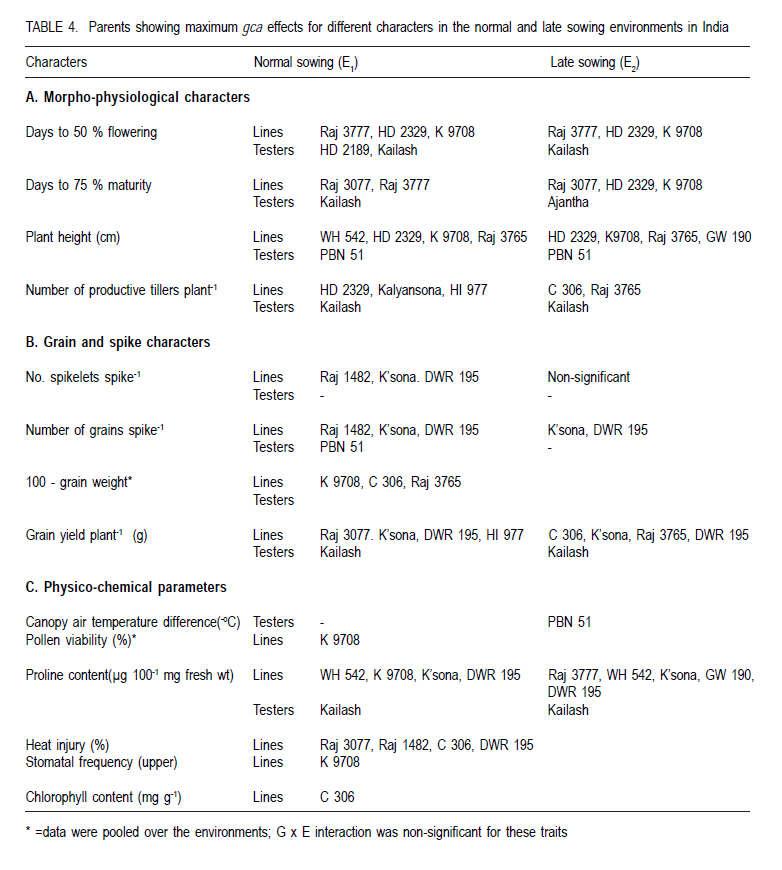
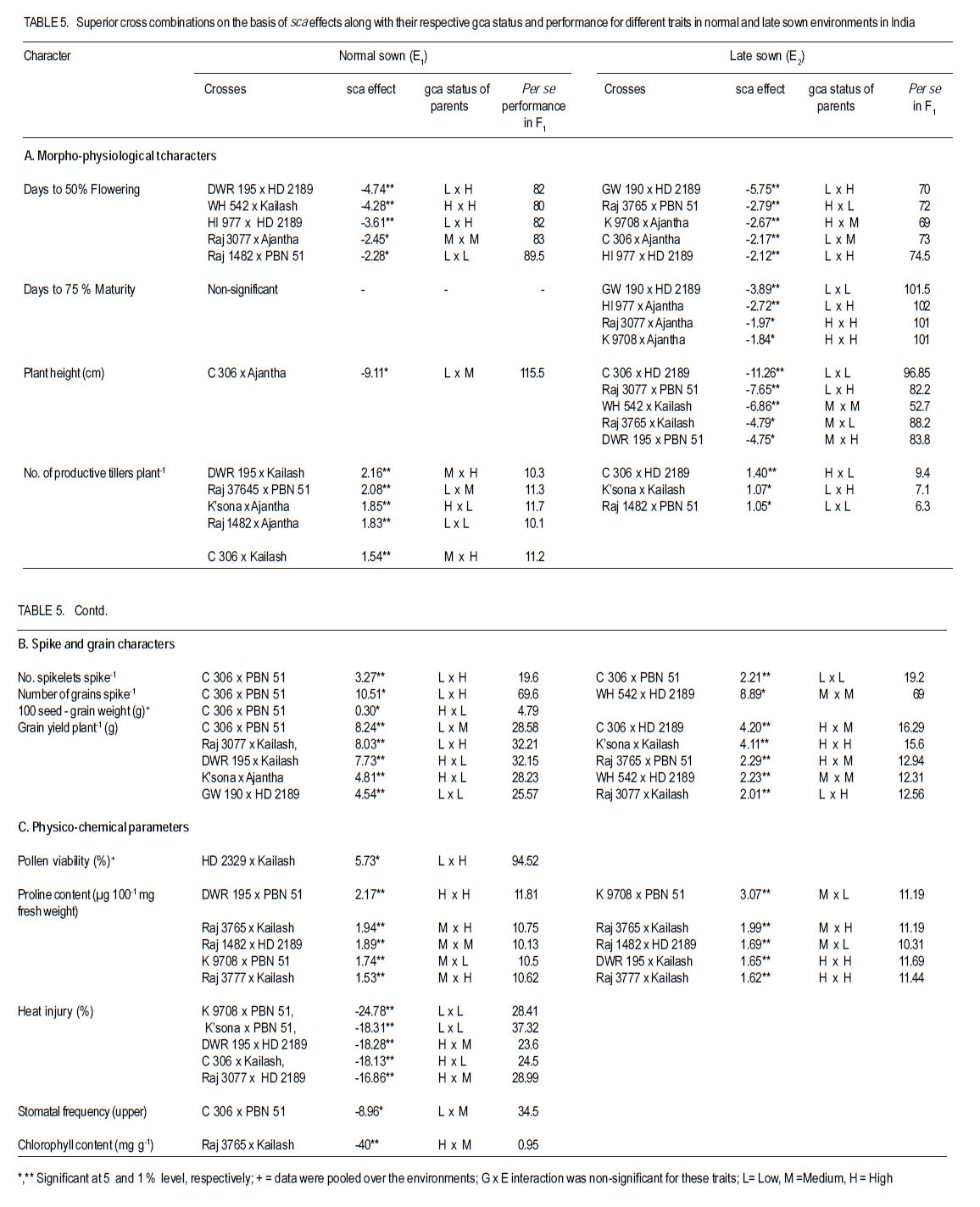
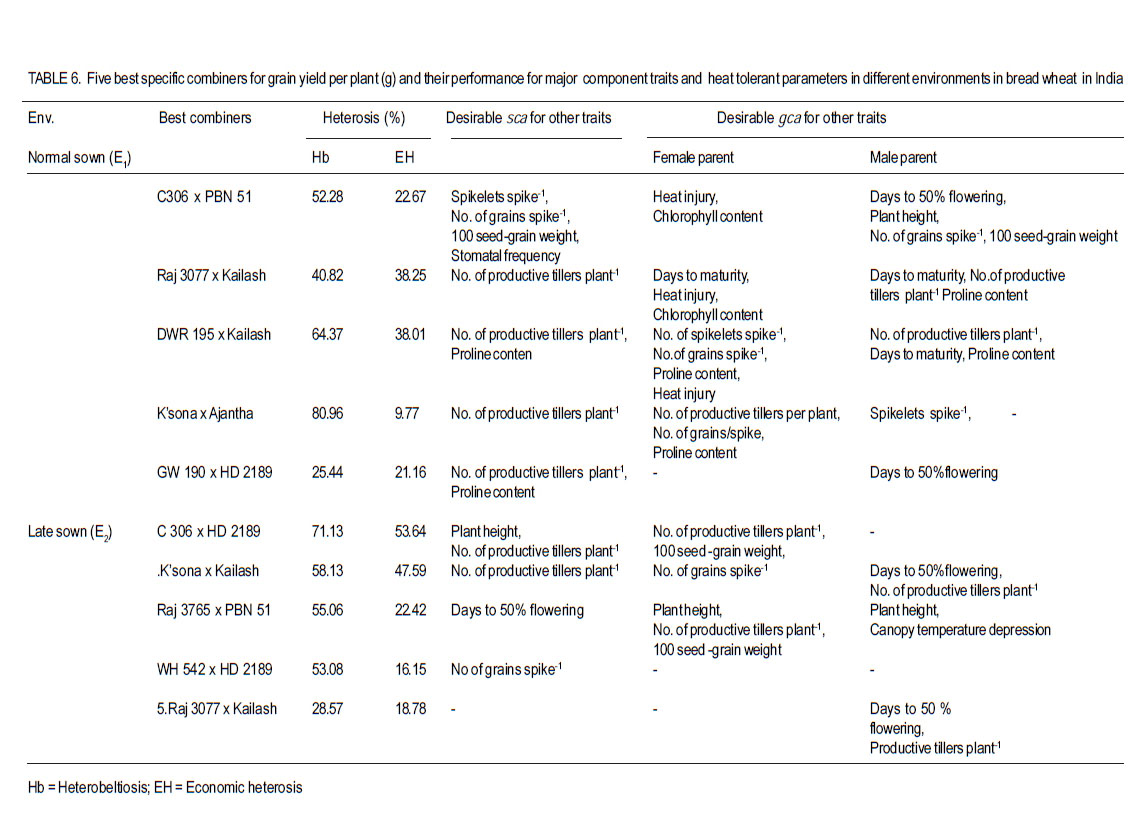
Where: = general mean, l = number of lines, t = number of testers, r = number of replications, X… = total of hybrid combination; Xi.. = total of ith line over testers and replications; X.j. = total of jth tester over lines and replications; and Xij. = ijth combination total over all replications. RESULTS L x T mating design for morpho-physiological, spike and grain and physico-chemical parameters revealed the presence of adequate genetic variability among parents and hybrids (Tables 1 and 2). Cumulative effect of gca and sca revealed the significance of both additive and non-additive gene actions in inheritance of these traits. High temperature and grain yield. Grain yield of parents and hybrids were reduced by more than 50% under the high temperature environment (E2) compared to normal sown (E1) (Table 3). Overall, the normal sown environment was the best for the yielding ability. The mean maximum temperature under E1 was 23.5 and 32.3 oC at reproductive and maturity stages, respectively. Whereas, under heat stressed (E2) environment, it was 31.8 and 36.4 oC at reproductive and maturity stages, respectively. gca effects Morpho-physiol-ogical characters. None of the parents was a good general combiner for all the characters under both environments (Table 4). Parental lines, viz., HD 2329, K 9708, Kailash, Raj 3077 and Raj 3777, were the best general combiners, exhibiting negative but significant gca for days to flowering, days to maturity and plant height. On the other hand, a positive significant gca was found for number of productive tillers per plant in E1. Of these, the first two parents also had desirable gca for days to flowering, days to maturity and plant height under E2. Spike and grain characters. Lines DWR 195 and Kalyansona were the best general combiners for most of the spike and grain characters under both the environments (Table 4). In respect to grain yield per plant, Kailash under both the situations, Raj 3077, HI 977 E1 and C 306 and Raj 3765 under E2 were promising parents. Physico-chemical parameters. None of the parent was an excellent combiner for all the traits studied (Table 4). However, tester PBN 51 showed significant and negative gca effects in E2 for CTD. Line K 9708 was a good combiner with significant and desirable gca effects for pollen viability, proline content (E1) and stomatal frequency on the upper side of the leaf. The parents, viz., Kailash, WH 542, Kalyansona and DWR 195, were good combiners for proline content under both conditions. On the other hand, Line C 306 showed maximum negative desirable gca effects for heat injury and chlorophyll content under both E1 and E2 . With negative significant gca effects, Raj 3077, Raj 1482 and DWR 195 were also good combiners for heat injury. Parents Raj 3777 and GW 190 were good combiners for proline content in E2 . sca effects Morpho-physiological characters. Five best cross combinations on the basis of significant and desirable sca effects are presented in Table 5. None of the cross combination showed desirable significant sca for both E1 and E2 for the traits studied. However, parental line DWR 195 in combination with HD 2189 (low x high gca) for days to 50% flowering (-4.74), with Kailash (medium x high gca) for number of productive tillers plant-1 (2.16) and C 306 x Ajantha (low x medium gca) for plant height (-9.11), had significant desirable sca effects with good performance (115.50) in E1. Whereas, under late sown situations (E2) crosses, viz., GW 190 x HD 2189 and K 9708 x Ajantha showed significant desirable sca effects with good per se for days to flowering (-5.75 and -3.89) and maturity (-2.67 and -1.84), respectively. The cross combination C 306 x HD 2189 for plant height (-11.26) with L x L gca and for number of productive tillers plant-1 (1.40) with H x L gca, displayed significant desirable sca effects with good per se performance in late sown situations (E2). Spike and grain characters. As for spike and grain traits, cross combination C 306 x PBN 51 was the best specific combiner since it exhibited maximum significant sca effect (3.27 and 2.21) in both conditions, respectively, for number of spikelets spike-1 ; but for other traits it showed significant sca effect only under E1 . The cross Raj 3077 x Kailash (low x high gca) had highly positive and significant sca effects and performance for grain yield under E1 (8.03 and 32.21) and E2 (2.01 and 12.56), respectively (Table 5). This cross also had high economic heterosis, 38.25 and 18.78, under both environments, respectively (Table 6). Whereas the other two hybrids, viz., DWR 195 x Kailash (7.73) and Kalyansona x Ajantha (4.81) with high x low gca, also depicted significant sca for grain yield in normal sown (E1) environment. On the other hand, crosses, viz., C 306 x HD 2189 (4.20) with high x medium gca and Kalyansona x Kailash (4.11) with high x high gca of parents showed significant sca effects for grain yield under late sown (E2) conditions. These two crosses also depicted maximum economic heterosis (53.64 & 47.59) under the late sown environment, respectively (Table 6). Physico-chemical parameters. Cross combination K 9708 x PBN 51 was superior for heat tolerance parameters, viz., for proline content with medium x low gca under E1 (1.74) and E2 (3.07) and for heat injury (-24.78) with combination of low x low gca (Table 5). In addition, the combinations viz., Raj 3765 x Kailash with medium x high gca (E1 and E2), Raj 1482 x HD 2189 with medium x medium (E1) and medium x low gca (E2), and Raj 3777 x Kailash with different level of gca status in different environments also showed high significant sca effect for proline content. The first combination was also a better specific combiner for chlorophyll content. As regards pollen viability on pooled analysis, HD 2329 x Kailash was the best specific combiner and the combination C 306 x PBN 51was the best for stomatal frequency on the upper side of the leaves. DISCUSSION Mean maximum temperatures around 23.5 oC at reproductive and 32.3 oC at maturity phase favour the growth and grain development in wheat. As the crop got exposed to higher ambient temperature at reproductive and maturity phases, significant reduction in productivity was realised (Tables 1 and 2). Under late sown environment (E2), drastic reduction in grain yield was likely due to rise in maximum temperature up to 31.8 oC at reproductive and 36.4 oC at maturity phases. Temperatures above 32 oC have been reported to reduce grain yield and grain weight (Wardlaw et al., 2002). In fact, Shoran et al. (2007) reported that each degree rise in ambient temperature reduces the yield by 3-4 %. Both general and specific combining ability effects played an important role in the control of morpho-physiological, spike and grain, and physico-chemical parameters of the genotypes studied. The specific effects being greater than the general effects (Tables 1 and 2). This suggests a prominent role of non-additive genetic effects, indicates that dominance and epistasis were major role in the expression of the traits. Similar results were reported in wheat by Sharma and Pawar (2000) and Kamaluddin et al. (2007). gca effects Morpho-physiological characters. Earliness seems to have favoured the plant to escape the losses due to terminal high temperature, while profuse productive tillering perhaps contributed to grain yield. Cultivar HD 2329 could be utilised for transfer of genes responsible for earliness and profuse tillering under terminal high temperature conditions. Parents such as HD 2329, K 9708 and Kailash may be potential components in the breeding programme intended to target short duration and stature genotypes. Profuse productive tillers is also an asset for wheat growing areas where high temperature stress at maturity affects overall crop yield. Early maturing and profuse productive tillering are desirable traits for high temperature tolerance Choudhary et al., 1996). Spike and grain characters. Lines C 306, Kalyansona and DWR 195 were the best combiners for grain yield under E2. This indicated presence of genes for heat tolerance (Table 5). Line Kalyansona and DWR 195, also good combiners for number of grains per spike, suggesting that number of grains per spike contributes to grain yield under heat stressed environments. These genetic resources are useful for improvement of heat tolerance in wheat breeding programmes. Physico-chemical parameters. Line K 9708 was exceptionally a good combiner for pollen viability, proline content and stomatal frequency on upper side, suggesting that good pollen viability status, more proline accumulation, along with lower stomatal frequency on upper side of leaves enhance the mechanism of heat tolerance in wheat. However, line C 306 showed low levels of heat injury and chlorophyll content that favour heat tolerance mechanism under terminal high temperature conditions. Deshpande and Nayeem (1999) also reported that parent C 306 was a good general combiner for heat injury. The negative gca for chlorophyll content, heat injury and stomatal frequency was affirmative for their importance in heat tolerance mechanism in wheat (Nayeem and Veer, 2000). sca effects Morpho-physiol-ogical characters. The estimates of sca effects were not consistent over the two environments viz., E1 and E2 (Table 5). An ideal genotype for E2 high temperature conditions is that which is early maturing, medium tall in height and has a good number of productive tillers per plant (Choudhary et al., 1996). Crosses, GW 190 x HD 2189 and K 9708 x Ajantha for maturity; C 306 x HD 2189 for plant height and number of productive tillers per plant, displayed desirable sca under E2. These crosses were derived from low x low, and high x low gca parents, thereby suggesting resorting to multiple crosses, followed by intermating (Singh and Paroda, 1987). Crosses involving at least one parent with high gca effect could produce good segregants, only if the additive genetic system present in the good general combiner and the complementary epistatic effects in the other act in the same direction to maximise the desirable plant attributes (Singh and Choudhary, 1995). In contrast to this, crosses showed significant desirable sca effects, even derived from low x low gca of parents. This is probably due to their wider genetic base and non-additive genetic effects. Spike and grain characters. The cross C 306 x PBN 51 was found best under normal sowing (E1), as it produced superior hybrids for yield and its component traits (Table 6). Raj 3077 x Kailash was found suitable for both conditions (E1 and E2). Hybrid WH 542 x HD 2189 was suitable under late sowing high temperature environment for yield and number of grains per spike (Table 6). These crosses which were derived from low x low, low x high and medium x medium gca parents, indicated the importance of non-additive genetic variation which can be exploited by multiple crosses, followed by intermating among desirable segregants. Kamaluddin et al. (2007) in wheat and Iqbal et al. (2007) in maize also reported crosses displaying high specific combining ability effects for seed weight and yield derived from parents with various types of general combining ability effects (high x low, low x low and medium x low). Majumdar and Bhowal (1968) earlier suggested that crossing low and high combiners in order to have desirable transgressive segregates might follow a high-low method of crop improvement for wheat. Physico-chemical parameters. Pollen viability was one of the desirable parameter for breeding heat tolerant varieties (Table 5). Cross HD 2329 x Kailash with low x high gca of parents may produce desirable segregants due to dominance and epistatic effects in F2. The combination of K 9708 x PBN 51 displayed desirable significant sca for proline content and heat injury, involving medium x low and low x low gca parents respectively. This suggests that it is preferable to resort to multiple crosses, followed by intermating.The superior cross combination involving low x low general combiners could result from over dominance and epitasis. However, crosses between medium x low indicated importances of non-additive genetic variation for the trait. In maize, Iqbal et al. (2007) also suggested that superior crosses involving high x low or medium x low combiner as parent could be explained on the basis of interaction between positive alleles from high/medium combiners and negative alleles from the low combiners as parent. The low stomatal frequency (upper) was also one of the desirable parameter while breeding for hest tolerant varieties. Cross C 306 x PBN 51 (8.96) with low x medium gca of parents displayed desirable sca for stomatal frequency (upper). On the other hand, for stomatal frequency (lower) none of the cross showed desirable significant sca. Nayeem and Veer (2000) also showed significant positive sca for chlorophyll content, and negative for stomatal frequency (upper) and heat injury. CONCLUSION The results signify the importance of exploitation of additive and non-additive genetic effects for attaining maximum improvement in grain yield along with heat tolerance. It appears, therefore, that single seed decent or the bulk method of handling segregating generations can also be used to utilise additive or additive x additive gene effects for developing better yielding cultivars suited to different environmental conditions, especially terminal heat stress. It is also suggested that high gca parents for grain yield and its component traits, and heat tolerant parameters, like C 306, Kalyansona, DWR 195, K 9708, Raj 3077, PBN 51, Kailash can be effectively utilised as a donor parent for the development of heat tolerant varieties. Improvement in heat injury and other heat tolerant parameters combined with high yield should be possible by resorting to biparental matting followed by recurrent selection or by selective diallel mating system. ACKNOWLEDGEMENT This study was supported by the Indian Council of Agricultural Research (ICAR), New Delhi, India. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11015t5.jpg] [cs11015t3.jpg] [cs11015t6.jpg] [cs11015t2.jpg] [cs11015t1.jpg] [cs11015t4.jpg] [cs11015e1.jpg] |
| |||||||||

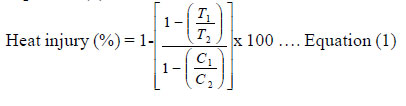
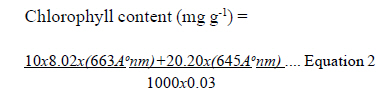
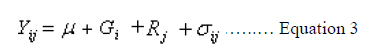
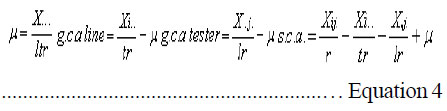
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}