 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 255-265 Seed storage proteins ARL2 and its variants from the APA locus of wild tepary bean 255 G40199 confers resistance to Acanthoscellides obtectus when expressed in common beans P.M. Kusolwa1 and J.R. Myers2
1Department of Crop Science and Production, Sokoine University of Agriculture, P. O. Box 3005, Chuo Kikuu
, Morogoro, Tanzania
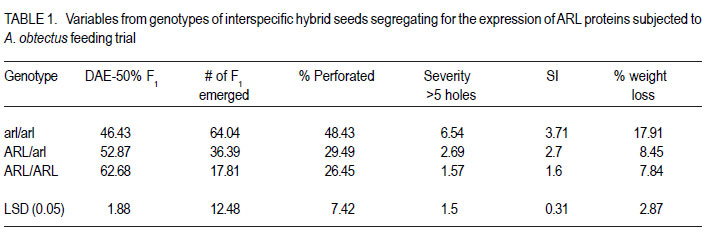
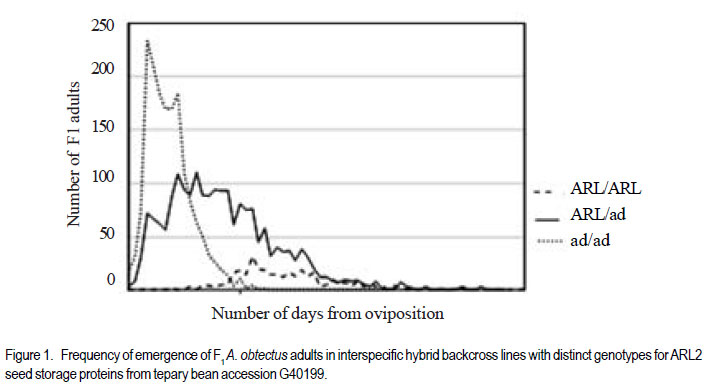
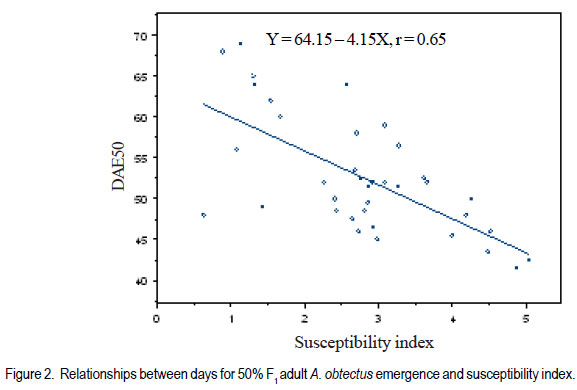
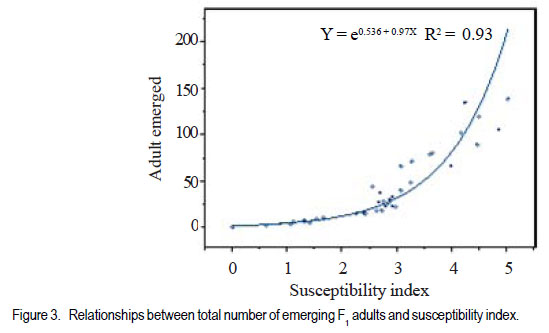
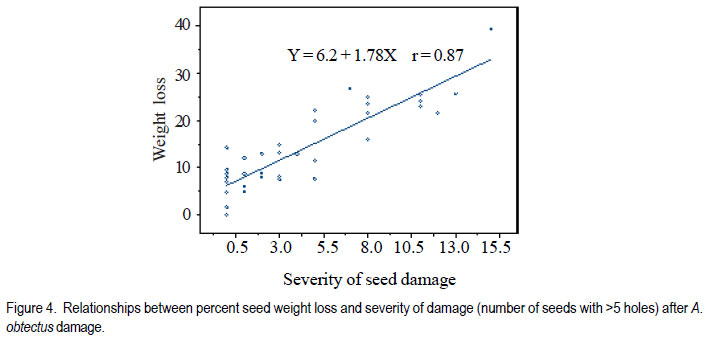
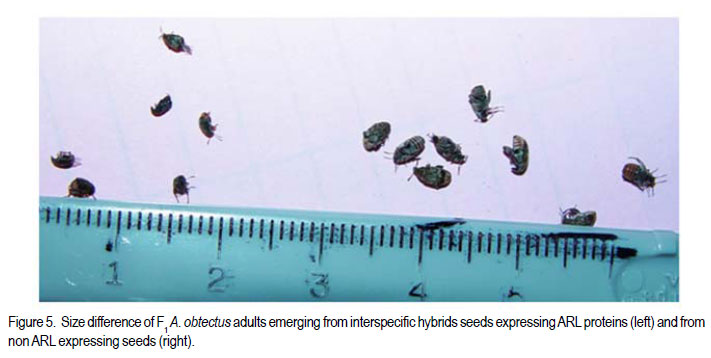
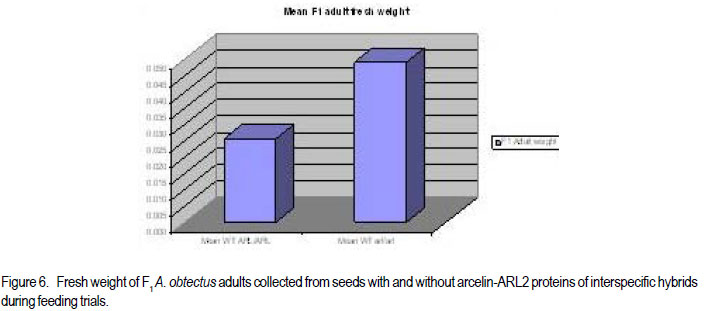
Code Number: cs11024 ABSTRACT Wild tepary beans (Phaseolus acutifolius) contain arcelins and arcelin-like (ARL2) proteins that are co-expressed with other proteins of the arcelin, α-amylase inhibitors and phytohemagglutinins (APA) locus. Arcelin and ARL2 protein from a wild tepary bean accession G40199 were transferred into common bean cultivars ICA Pijao and Rojo. Inbred backcross lines were developed and selected for segregation of the proteins at BC2F2:3 generations. Seeds from homozygous, segregating lines expressing the arcelin and ARL2, plus those without the proteins, were used for screening against resistance to A. obtectus. Significant delay of 63 days for 50% adult emergence and reduced number of emerged F1 of A. obtectus adults were observed in backcross lines with homozygous expression of arcelin and ARL2 tepary bean proteins. Furthermore, a reduction in size and weight of emerged adult insects to almost half was observed. This work demonstrates the superior resistance common bean backcross lines to A. obtectus conferred by the presence of the APA proteins introgressed from tepary bean. Key Words: Arcelin, Phaseolus acutifolius RÉSUMÉ Les haricots sauvages tepary (Phaseolus acutifolius) contiennent des protéines arcelines et les protéines semblables aux arcelines (ARL2) co-exprimées avec d’autres protéines arcelines, tels les inhibiteurs α-amylase and les locus phytohémagglutinines (APA). Les protéines arcelines et ARL2 provenant des accessions de haricots sauvages tepary G40199 étaient transférés dans les cultivars du haricot commun ICA Pijao et Rojo. Des lignées issues de croisement en retour étaient développées et sélectionnées pour la ségrégation des protéines aux générations BC2F2:3. Des graines des lignées homozygotes ségrégantes exprimant les protéines arcelines et ARL2, plus celles sans protéines, étaient utilisées pour la sélection contre la résistance au A. obtectus. Un retard significatif de 63 jours pour 50% de l’émergence des adultes et un nombre réduit de F1 émergées des adultes A. obtectus étaient observés dans le croisement reour avec expression des homozygotes des protéines arcelines et ARL2 des haricots tepary. En outre, une réduction jusqu’à la moitié en taille et poids des insects apparus était observée. Cette étude démontre la résistance supérieure des lignées de croisement en retour au A. obtectus conférée par la présence de protéines APA introgressées du haricot tepary. Mots cles: Arceline, Phaseolus acutifolius INTRODUCTION In the tropics, dry bean (Phaseolus vulgaris L.) production is constrained by several biotic and abiotic factors. Some of the major biotic constraints involve post-harvest losses caused by two major bruchid species Acanthoscellides obtectus and Zabrotes subfasciatus. The bean weevil, A. obtectus is a predominant post-harvest pest in all tropical highland bean producing regions, attacking beans in the field and in storage. The pest causes quality degradation and poor germination of damaged seeds. In an effort to find sources of resistance to these pests, evaluation of bean germplasm has identified seed proteins of the lectin-like family, called arcelins, found in wild common beans (Osborn et al., 1988) associated with resistance to bruchids. Highest level of resistance to Z. subfasciatus was described to be found in in wild common bean accessions and is associated with the expression of arcelin 5, arcelin1, arcelin2 and arcelin 4 in order of effectiveness (Cardona et al., 1990; Kornegay et al., 1993). Breeding strategies to improve resistance of commercial cultivars of dry bean have involved backcrossing method to introgression of arcelin proteins from wild genotypes of common beans (Osborn et al., 1986; Osborn et al., 1988; Cardona et al., 1990; Kornegay and Cardona, 1991; Kornegay et al., 1993; Misangu, 1997). High levels of resistance to two major bruchid species were demonstrated in tepary bean genotypes by Shade et al. (1987) and recommended for use as potential source of resistance. Further reports demonstrated that lectins present in some tepary bean accessions conferred high resistance to the bean weevil Acanthocellides obtectus (Pratt et al., 1984; 1990). Arcelin and arcelin-like proteins have been reported in cultivated accessions of tepary bean P. acutifolius (Mirkov et al., 1994; Gerhardt et al., 2000), but their resistance to A. obtectus was not characterised. Arcelins and ARL proteins present in tepary beans are part of the lectin-like proteins which may exist as different alleles and variants (Lioi et al., 2003; 2006). Since different variants of arcelins or arcelin-like proteins may confer different levels of resistance to bruchids, a wild tepary bean accession G40199 from CIAT and preliminary studies to use it as a resistant source to bruchids was proposed by Myers et al. (2001). Detailed characterisation of total seed storage proteins from this accession demonstrated that it contains a complete APA locus of the lectin-like proteins. This accession demonstrated expression of arcelin, arcelin-like (ARL2) and phytohemagglutinins, lectin like proteins specific to tepary beans only. It was hypothesised that the transfer of these proteins into common bean cultivars may improve resistance to A. obtectus. These proteins were transferred by interspecific hybridisation via embryo rescue (Mbogo et al., 2009). This work is aimed at evaluating the association of the introgressed APA genes from G40199 x common bean interspecific backcross lines selected for segregating locus with ARL2 seed proteins. MATERIALS AND METHODS Plant materials and genotype identification. A population of interspecific hybrid backcross families was used to screen for A. obtectus resistance. Interspecific hybrid lines from BC2F3 seeds from a cross between ICA Pijao and tepary bean accession G40199 were used in the bruchid screening trials. A total of eleven single plants from each backcross family was randomly selected based on genomic DNA analysis for the presence of ARL2, á-AI and PHA by PCR amplification. However, it was not possible to separate homozygous from heterozygous plants for the genes at this generation. Therefore, seeds from ARL2+ single plant selection (SPS) were harvested and used to progeny test the BC2F2:3 by protein gel electrophoresis to identify individual lines that do not segregate for the 33KDa tepary protein corresponding to the APA proteins. Total seed proteins were analysed on eleven seeds from each individual backcross lines using 15% tris-HCl SDS –PAGE. Three genotypes were identified: homozygous (ARL/ARL, 26 lines), heterozygous (ARL/arl, 64 lines), and homozygous null (arl/arl, 36 lines) based on the 33kDa storage proeins. Four samples containing bean seeds from each cultivars Rojo, ICA Pijao were used as susceptible checks for parents, while four vials with accession G40199 seeds were included as resistant control of the parent wild tepary bean. Screening for resistance to A. obtectus among tepary bean hybrids. A spontaneous colony of A. obtectus was obtained from bean stocks in the Oregon State University (OSU) dry bean breeding programme. The colony was multiplied and maintained in susceptible Rojo and Pinto bean seeds at OSU to obtain enough adult insects for inoculation of experimental materials. A total of fifteen non-sexed A. obtectus adults were placed into the glass vials containing 30 seeds from interspecific backcross lines with ICA Pijao as recurrent parents. Glass vials were loosely closed to allow aeration but prevent escape of adult insects placed into incubation trays, kept undisturbed for 12 days at 25 ± 3°C. After 12 days, the number of laid eggs was recorded using a magnifying lens and then adults were removed. In cases where no laid eggs were visible, samples were re-inoculated with fresh adults. Vials were placed back into the incubation tray and left undisturbed but monitored until the first adult emerged. Powdery/flour appearance on the surface of the beans was noted to be sure of the larvae penetration into the seeds and identify for possible escapes. Following the observation of the first emergence of F1 adults, each vial was inspected daily and emerging F1 adults were counted and removed to avoid laying of new eggs for a period of 72 days after inoculation of adults. Further observation was continued to 100 days in some lines that showed a delayed emergence of F1 adults, especially in samples of the parent G40199 tepary beans and some of the interspecific hybrids. In an alternative attempt, seed coat from G40199 seeds was removed by scratching to verify if the hard seed coat was or not a factor for resistance to development of A. obtectus larvae in these seeds. Data collected included the total number of F1 adults emerging after inoculation, number of days for first adult emergence (DAE), number of days for 50% of total F1 adults emerged and susceptibility index (SI) calculated as the natural log. Other variables evaluated were the number of perforated seeds, severity of damage expressed by the number of seeds with 5 or more holes and percent seed weight loss. Frequency of adult emergence was determined by the total number of F1 adults emerged per day for the period of 72 days. During the experiment, samples of emerging adults were collected from vials of beans with and without the ARL2/arcelin proteins and weight of 10 adults taken for each group. Variables were subjected to analysis of variance, regression analysis for correlation of the novel seed protein to resistance Acanthocelides obtectus using SAS 9.1.3 and SPLUS 6.2 programmes (for graphics). RESULTS Delay for adult F1 emergence. Significant delay for 50% F1 adult emergence was observed in interspecific hybrid seeds containing ARL2 proteins (P <0.0001). Adult emergence was delayed to an average of 63 days for adults emerging from seeds that were homozygous for the expression of ARL2/arcelin seed proteins inherited from tepary bean G40199. While in the absence of the proteins, insect emergence was observed earlier (Table 1). Reduced number of emerging F1 adult bruchids. A significantly lowest number of emerging F1 adults was observed in genotypes that were homozygous for the presence of ARL2 proteins (Table 1). Overall, only 18 F1 adults were observed in these lines for a period of 72 days following oviposition. The highest number was observed in seed lots that did not express the ARL2 proteins. In addition, the frequency of F1 adult emergence was low in seeds containing the ARL2 proteins (Fig. 1). A higher number of F1 emerged early in susceptible materials without the ARL2 seed protein, followed by a fast drop a few days after the first adult emergence. While in homozygous ARL2 containing seeds, low frequency with fewer number of adult F1 emerged late and slowly over long time implying a significant retardation of the reproduction cycle for the insect in this bean stock. The peaks for frequency of adult emergence corresponded to the mean number of days for 50% of F1 adult emerged in each of the segregating /ARL2 genotypes (Fig. 1). Seed damage. Significant differences occured in percentage perforated/ damaged of seeds (P <0.0001) as a result of larvae feeding the activities leaving holes and or dusty beans (Table 1). Bean seed lots containing arcelin and ARL2 proteins were less damaged with lowest number of perforated seeds. Very few seeds with 5 or more holes were observed in these seed lots, while up to 7 holes per seed could be observed in samples without this storage protein. Susceptibility index. Susceptibility index (SI) was significantly lowest with a value of 1.6 for homozygous lines containing the ARL2 proteins, while the highest at 3.71 for backcross lines with no ARL2/arcelin proteins (Table 1). The SI was negatively correlated with number of days for F1 adult emergence, whereby ARL2 containing seeds with higher DAE-50% and low SI (Fig. 2). Susceptibility index also demonstrated exponential relationships with the number of emerging F1 adults indicating that more susceptible seeds (high SI) had exponential increase in emergence of new adults (Fig. 3). Homozygous backcross lines for arcelin and ARL2 proteins had less emerging adults with low SI, while in the absence of the proteins and higher SI resulted into exponential increase in total number of emerging adults and total damaged seeds. Reduction of seed weight. Significant reduction in seed weight was observed as a result of larvae feeding on the seeds (P<0.0014). Genotypes from interspecific backcross families containing arcelin and ARL2 proteins had the lowest mean seed weight loss (7.84%) compared with 17.91% in genotypes within the same backcross family, but without the protein (Table 1). Seeds from segregating genotypes had no significant difference for seed weight loss when compared with homozygous ARL2 containing seeds. Seed weight loss was highly correlated with severity of damage (Fig. 4). Bean seed lots with severe perforation (more than 5 holes seed-1) demonstrated high seed weight loss. Bean samples without ARL proteins had the highest number of severely damaged seeds, with more than 5 holes and thus suffered higher seed weight loss. Seed weight loss was also highly correlated to total number of perforated seeds. No adults of A. obtectus that emerged from seed samples of the parent tepary bean G40199 after a period of 120 days. Larvae penetration into the seeds was clearly visible as powdery/ floury appearance of the seeds and no larvae were observed to remain outside the seeds (not presented). But no adult emerged after another 90 days of observation following the oviposition. Similarly, no F1 adults emerged from some of the backcross lines with homozygous presence of ARL2 of the APA proteins (ARL/ARL). These interspecific lines were kept for further screening and seed increase. Weight and size of F1 A. obtectus adults Reduced fresh weight of F1 adults. Significant differences in size of emerging adults were observed (Fig. 5). Earlier emerging F1 adults from ICA Pijao, Rojo and backcross lines with null expression of arcelin and ARL2 proteins were consistently larger in size than the emerging adults from seeds with ARL2 of the APA proteins. The mean live weight of 10 F1 adults emerging from arcelin plus ARL2 seeds were significantly low with mean insect weight almost twice lower than the adults emerging from non ARL protein seeds (Fig. 6). DISCUSSION The observed resistance to A. obtectus was consistently associated with the expression of a 33kDa seed storage lectin, like proteins among interspecific hybrids. This is a major Lectin-Like Protein (LLP) profile that cosegregates with the expression of arcelin and ARL2 genes in G40199 and interspecific hybrids as observed in a previous study (Mbogo et al., 2009). Homozygous lines demonstrated higher resistance compared to the segregating genotypes in the two backcross populations. However, there were variations in number of F1adults that emerged from homozygous ARL/ ARL containing seeds and delay in 50% adult emergence among individual backcross lines. If the resistance is highly associated with the presence of the 33kDa and the other linked subunits of LLP, then a variation in concentration, composition and conformation of the proteins may exist among the seeds of genotypes expressing the ARL2 proteins. These variations were some of the factors contributing to variations in means for variables observed during the screening trial. Some lines expressing the 33kDa proteins profiles were not at all damaged during feeding trial, and were reinoculated for the second time then only few weak F1 adults emerged late. The delay in adult emergence observed among interspecific hybrids containing homozygous expression of the 33kDa protein profiles (Fig. 1) were remarkably high compared with susceptible cultivars Rojo and ICA Pijao used in the hybridisation. A mean delay for first adult emergence of 62.8 days (two months after infestation) among interspecific genotypes containing the protein is a significant factor for storage in tropical bean producing countries. Some of the backcross interspecific hybrids lines had as high as 69 days delay in emergence of first adult (Fig. 1). This means significant damage may occur very late during storage, farmers may be able to avoid this damage with minimum integration of pest management strategies like cultural, use of plant product (Schhonhoven and Cardona, 1986; Mazzonetto and Vendramim, 2003) that will further prolong the DAE for new adults and increase adult mortality of bruchids. In addition to delayed emergence, emerging adults from these bean genotypes were fewer, small size and weak. Full resistance to A. obtectus was only observed in samples of wild tepary bean G40199 where no single F1 adult emerged from either beans with seed coat removed or beans with intact seed coat. The total absence of damaged seeds in the parent G40199 indicates that the 33kDa lectin-like proteins in this accession may not be the only factor responsible for its antibiosis. Alternatively, the concentration of LLPs in this accession may be significantly high to inhibit any development of larvae inside the seed. The 33kDa protein in G40199 was observed as the major protein almost replacing phaseolin content in the total seed storage protein (Kusolwa, 2007). Backcross interspecific hybrid lines with the 33kDa proteins that demonstrated high antibiosis to the bruchids, may have a relative higher expression of the protein to the level comparable to that of G40199. Genomics and proteomics studies from Mbogo et al. (2009) and Kusolwa and Myers (2010) demonstrated that the accession G40199 contain the APA locus with variants arcelins and other APA locus proteins are expressed in seeds of this genotype. The tightly linked complex APA locus was transferred into common bean by interspecific hybridisation demonstrating the presence of expression of arcelins alpha amylase inhibitors and phytohemagglutinins. It was, therefore, expected that the expression of APA locus in interspecific hybrids will confer the same level of resistance to bruchid similar to G40199. However, bruchid screening from interspecific hybrids shows that the introgressed APA locus highly increased resistance of common bean genotypes to A. obtectus though not to same level of resistant as of the parent source tepary bean G40199. These results indicate that the APA locus containing arcelin and ARL2 proteins is not the only factor associated with full resistance in G40199. Tepary beans are known to contain high quantities of other antinutritional proteins and other compounds (Idouraine et al., 1992; Magdi et al., 2003; Campos et al., 2004) that may play an important role in antibiosis to bruchids. These proteins and probably other factors yet to be identified were likely not transferred into common bean by interspecific hybridisation or might have been lost during backcrossing and bridge parent hybridisation process. Chromosome elimination and loss of quantitative traits among interspecific hybrids in common bean x tepary bean crosses are known (Rabakoarihanta et al., 1979; Federici and Waines 1989; Mejia Jimenez et al., 1994; Muñoz et al., 2004). When the total seed storage proteins from interspecific hybrids and G40199 were compared by SDSPAGE profiles, modification of the 26kDa and 28kDa were observed (Kusolwa, 2007). These may be part of factors that modify the composition of protein subunits of the APA locus in the interspecific hybrids resulting into difference in levels of resistance. To minimize this problem, we developed in bred congruity backcross lines of interspecific hybrids to G40199 containing two doses from P. acutifolius and P. vulgaris (ICA Pijao), followed by two selfing generations. The reduction of fresh F1 adult weight in insects emerging from seeds containing homozygous expression of the ARL2 proteins is an important supporting evidence that this protein is highly associated with resistance to A. obtectus. Poor digestibility of arcelin proteins by insect’s larvae enzymes was observed in arcelin containing beans (Minney et al., 1990; Goossens et al., 2000; Paes et al., 2000) this might have contributed to adult size and weight reduction of emerging adults. The arcelin and ARL2 proteins introgressed into interspecific hybrids may have contributed to the disruption of the digestive system of the larvae thus affecting its development. The weak-emerging adults may have suffered other physiological disabilities that contributed to the total delay in instar development, and possible reproductive disorders may also be expected. Consequently, the subsequent insect generations may experience low fecundity causing further weakening the entire bruchid colony in a given bean stock containing this ARL2 and its variants from the APA proteins. Significant reduction in seed damage demonstrated by reduced seed weight loss and low number of severely perforated seeds with 5 or more holes was highly associated with these introgressed proteins among the backcross families. Low seed damage was highly correlated with the reduced adult F1 fresh weight, low susceptibility index, and reduced number of emerging F1 adults. Although the full resistance observed in G40199 was not transferred into two common bean lines by interspecific hybridisation, the level of resistance reported in homozygous lines for the expression of ARL2 is highly significant compared with the susceptible parent common bean cultivars Rojo and ICA Pijao. Delayed F1 adult emergence, reduced number of F1 adults, and reduction in size and weight of adults are important factors for bruchid resistance common bean cultivars. Bruchid screening confirmed the stability of the introgressed ARL2 of the APA locus proteins into common beans as suggested by the genomic analysis for expression of APA genes in backcross lines (Kusolwa and Myers, 2010). To our knowledge, this is the first work that describes the transfer, stable introgression into common bean cultivars and characterisation of arcelins and ARL2 proteins in tepary bean accession G40199. This highly resistant accession has been used by other researchers to study resistance to bruchid species and efforts to make interspecific hybrids with common beans have been attempted (Mejia Jimenez et al., 2002; Cardona et al., 2005). However, no stable self reproductive plants have been developed with resistance to A. obtectus. Prior to this work, characterisation of antibiosis factors of the lectinlike protein variants in this accession and their association to resistance to A. obtectus was not documented. This is the second stable abiotic resistance factor transferred from tepary bean to common bean cultivars in addition to resistance to common bean bacterial blight (Xanthomonas campestris pv. phaseoli) ( Singh and Muñoz, 1999). REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11024f5.jpg] [cs11024t1.jpg] [cs11024f6.jpg] [cs11024f1.jpg] [cs11024f2.jpg] [cs11024f4.jpg] [cs11024f3.jpg] |
| |||||||||

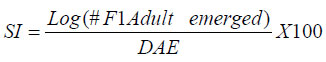
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}