 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 267-275 Inheritance of resistance to angular leaf spot in yellow bean. J.W. Muthomi, K.K. Muimui1 and P.M. Kimani
Department of Plant Science and Crop Protection, University of Nairobi, P. O. Box 30197-00100,
Nairobi, Kenya
1Misamfu Regional Research Station, P. O. Box 410055, Kasama, Zambia
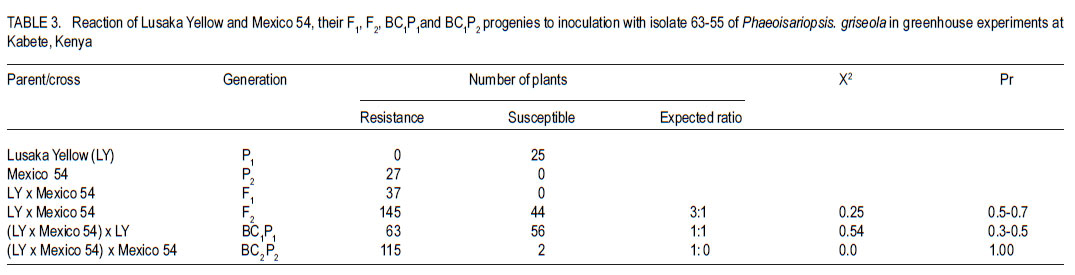
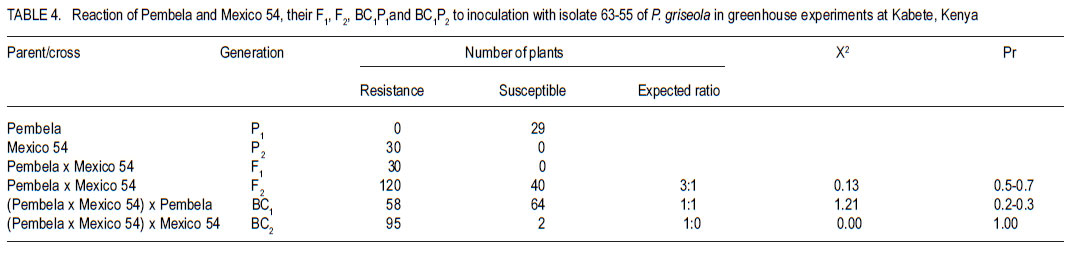
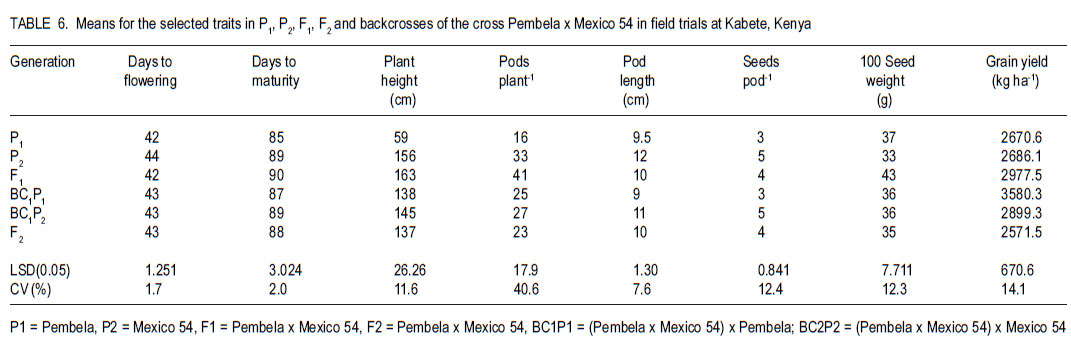
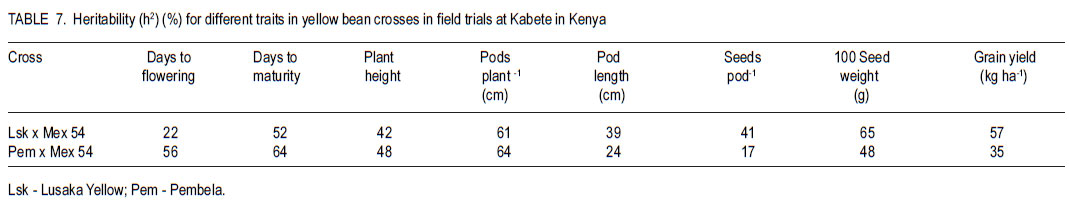
Code Number: cs11025 ABSTRACT Angular leaf spot (Phaeoisariopsis griseola (Sacc) is an important disease of common bean (Phaseolus vulgaris L.) in most parts of Africa, causing yield losses of 40-80%. This study was carried out to determine the inheritance of resistance to angular leaf spot in yellow beans. Biparental crosses were done between susceptible yellow bean genotypes and angular leaf spot resistant parents to generate F1, F2 and backcrosses. Resistance was evaluated in a screen house and field after inoculation with Phaeoisariopsis griseola. The F1 were resistant, indicating that resistance was dominant. There was no significant deviation from the expected 3:1 ratio for resistant to susceptible in the F2 population, confirming that resistance to angular leaf spot was both monogenic and dominant. The backcross to Lusaka Yellow showed a 1:1 segregation ratio, while the backcrosses to Mexico 54 were all resistant. Pembela was susceptible to angular leaf spot, while the F1 were resistant. The ratio of 3:1 represents resistant : susceptible F2 populations. All backcrosses to Mexico 54 were resistant, confirming that resistance to angular leaf spot in Mexico 54 is controlled by a single dominant gene. The results of the study showed that resistance to angular leaf spot in yellow beans is governed by a single dominant gene. This can be used to improve the local landraces by incorporating angular leaf spot resistance and high yield traits. Key Words: Monogenic, Phaeoisariopsis griseola, Phaseolus vulgaris RÉSUMÉ La tâche agulaire des feuilles (Phaeoisariopsis griseola (Sacc) est une maladie importante du haricot commun (Phaseolus vulgaris L.) dans la plus part de regions africaines, causant des pertes de rendement d’environ 4080%. Cette étude était conduite pour déterminer l’acquisition de la résistance des haricots jaunes à la maladie de tâche angulaire. Les croisements biparentaux étaient effectués entre les génotypes du haricot jaune susceptible et les parents résistants à la génération F1, F2 ainsi que de croisements récurrents. La résistance était évaluée en serre et au champ après inoculation avec Phaeoisariopsis griseola. Les F1 étaient résistantes, indicant que la résistance était dominante. Il n’avait aucune déviation significative du rapport attendu 3:1 pour la résistance, au susceptible dans les populations F2, confirmant que la résistance à la tâche angulaire de feuilles était monogénique et dominant. Les produits du croisement en retour de Lusaka Yellow a montré un rapport de ségrégation 1:1, pendant que ceux de Mexico 54 étaient tous résistants. Pempela était susceptible à la tâche angulaire, alors que les F1 étaient résistantes. Les progénies F2 ségrégées en un rapport 3:1 pour résistant au susceptible et le backcross (croisement en retour) au Pempela avait donné un rapport 1:1 pour résistant au susceptible. Tous les produits du croisement en retour du Mexico 54 étaient résistants, confirmant que la résistance à la tâche angulaire des feuilles dans Mexico 54 est controllée par un seul gène dominant. Les résultats de cette étude ont montré que la résistance à la tâche angulaire dans les haricots jaunes est gouvernée par un seul gène dominant. Ceci peut être utilisé pour améliorer les variétés locales indigènes par incorporation de la résistance à la tâche angulaire des feuilles et les traits à rendement élevé. Mots Clés: Monogénique, Phaeoisariopsis griseola, Phaseolus vulgaris INTRODUCTION Common bean (Phaseolus vulgaris L.) is the most widely grown legume and it is the second most important source of dietary protein in Eastern and Southern Africa (Pachico, 1993). Major seed classes grown include red mottled, large red kidney, small red, yellow, navy, purples, black and sugars (Wortmann et al., 1998). However, yellow bean cultivars are susceptible to diseases such as angular leaf spot (Phaeoisariopsis griseola) resulting in low yields. In Africa, angular leaf spot is considered to be the most widely spread and economically important disease of beans (Wortmann and Allen, 1994). The disease significantly reduces the number of seeds per pod as well as seed weight. It also leads to premature defoliation, shrivelled pods and shrunken seeds (Santos - Filho et al., 1976). Yield losses can reach 80% under severe conditions of infection (Schwartz et al., 1981). Management of angular leaf spot and other foliar diseases of beans is constrained by the high cost of chemicals, the ability of the pathogen to survive in plant debris for a long time and land un availability for crop rotation (Deeksha et al., 2009). The current focus is to develop varieties that have multiple-constraint resistance (Miklas et al., 2002; Sharma et al., 2007). Therefore, breeding programmes aim at pyramiding or accumulating several resistance sources in a variety as a way of developing broad and durable resistance in common bean in general, and in preferred bean classes in particular. To realise this goal, it is important to identify sources of resistance to the diseases, determine their mode of inheritance and deploy the resistance genes in order to improve the preferred but low yielding types in the market classes. Diverse sources of resistance to angular leaf spot in bean genotypes have been reported (Correa et al., 1989; Beebe and Pastor-Corrales, 1991). Examples of resistant cultivars include A 75, A140, A152, A175, A229, BAT 76, BAT 431, BAT 1432, BAT 1458 and G5686 (CIAT,1984). Regagnin et al. (2005) found angular leaf spot resistance in AND 277, while Nietsche et al., (2000) found it in Cornell 49-242. Mahuku et al. (2004) found resistance in G 10474. CIAT (2003) reported resistance in Mexico 54. Sources of resistance reported from Africa include GLP 24, GLP X-92, GLP - 806 and GLP 77 (CIAT, 1984). Resistance to various diseases is monogenically determined, but cases of duplicate, complementary and other interactions have been reported (Singh and Muñoz, 1999). The objective of this study was to develop segregating populations of yellow beans with resistance to angular leaf spot and determine the inheritance of the resistance. MATERIALS AND METHODS Generation of experimental populations. The experimental materials consisted of angular leaf spot susceptible Zambian varieties (Lusaka Yellow and Pembela), resistant line (Mexico 54) and their crosses (F1, F2, backcrosses BC1P1 and BC1P2). The progeny derived from backcrossing F1 to the female parent was designated BC1P1 and those from the backcrossing to the male parent as BC1P2. Lusaka Yellow and Pembela, which are widely grown and preferred in Zambia, but are susceptible to angular leaf spot and common bacterial blight, were used as females (Table 1); while the source of resistance was Mexico 54 whose seed was obtained from the Regional Bean Programme at the University of Nairobi. The two crosses were Lusaka Yellow X Mexico 54 and Pembela X Mexico 54. Emasculation and pollination was done either early in the morning or evening as described by Tumwesigye (1988). Buds, which were plump, showing colour and would open the following day were chosen as the female flowers. After emasculation of the female flower, pollination was done immediately using the rubbing or hooking method (Buishand, 1956; CIAT, 1987). Freshly opened flowers were chosen from donor parents to provide pollen. After crossing, a cotton thread and a tag labelled with the pedigree of the cross was tied loosely on the flower stalk. At maturity, the pods were harvested together with their identification tags. These were sun-dried and threshed to give F1 seed. Part of the F1 seed from each cross was sown in the screen house to produce F2 seeds and also backcrossed to both parents. Isolation of Phaeoisariopsis griseola and plant inoculation. Isolation of angular leaf spot was made from lesions of naturally infected bean leaves showing fungal sporulation. In the case of non-sporulating lesions, the fungus was induced to sporulate by incubating the infected tissues in moist chambers. Small pieces of infected tissues were surface sterilised with 3% (v/v) sodium hypochlorite for 5 minutes and plated on bean leaf decoction agar (BLDA, Karanja et al. 1994). The emerging fungal growth was subcultured on fresh BLDA medium. The inoculum was harvested in sterile distilled water and the suspension was filtered through a triple layer of muslin and adjusted to 2x106 conidia ml-1 using a haemocytometer. Three weeks old bean seedlings which were planted in inoculation chambers maintained at high humidity of over 90% 24 hours before inoculation. High humidity in the inoculation chambers was ensured by lining with polythene sheets and wet absorbent clothing. Using a hand atomiser, the plants were inoculated mainly on the abaxial side of the first primary and trifoliate simple leaves at a distance of 10-15 cm until run off. Controls consisted of plants treated with sterile distilled water without the pathogen inoculum. A repeat inoculation was done at the start of flowering. After inoculation, the plants were maintained under high relative humidity to enhance the development of angular leaf spot symptoms. Green house and field experiments lay out. Treatments consisted of angular leaf spot susceptible parents (Lusaka yellow and Pembela), resistant parent (Mexico 54) and their F1, F2 and backcrosses. The F1 and F2 crosses were Lusaka yellow x Mexico 54 and Pembela x Mexico 54 while the backcrosses were (Lusaka yellow x Mexico 54) x Lusaka yellow, (Lusaka yellow x Mexico 54) x Mexico 54, (Pembela x Mexico 54) x Pembela and (Pembela x Mexico 54) x Mexico 54. In the green house experiments, three to four seeds of each of the lines were planted in each of three pots per line and laid out in a completely randomised design with three replications. Sisal ropes were used to support the plants that showed some climbing tendency. Field experiments were conducted at the Faculty of Agriculture field station, University of Nairobi, located at latitude 11o 4’ 20" S and longitude 36o 45’ E with an altitude of 1820 metres above sea level. The site has an average annual rainfall of 1046 mm, mean maximum temperatures is 23o C and a minimum of 12o C. The soils are deep, well drained, dark-reddish brown friable clays. Three metre rows of each of the parents, F1, F2 and the backcrosses were planted per plot at a spacing of 50 cm apart and 15 cm within rows. The experiment was laid out in a randomised complete block design and replicated three times. Fertilisation was done at planting by application of di-ammonium Phosphate (DAP - 18% N and 46% P205) at the rate of 150 kg ha -1. Bean lines showing climbing growth habits were supported with sticks. Angular leaf spot severity, plant height, days to flowering and maturity and yield data were recorded for each genotype. Disease severity assessment was initiated at onset of symptoms while plant height was assessed at pod set. The yield components assessed included the number of pods per plant, pod length and seed yield. At physiological maturity, each genotype was harvested, air-dried and threshed separately to determine the number of seeds per pod, number of seeds per plant, 100 seed weight and total seed weight. Assessment of angular leaf spot severity.Angular leaf spot assessment was initiated at the onset of disease symptoms and each genotype was rated as resistant or susceptible based on a 1-9 assessment scale (van Schoonhoven and Pastor-Corrales, 1987) (Table 2). Scoring for the disease was done on three trifoliate leaves selected at the bottom, middle and top of each plant. The mean disease score for all the plants assessed was calculated for each genotype. Genotypes showing a disease severity rating of 1-3 were regarded as resistant, 4-6 as intermediate and 7-9 as susceptible. Statistical analysis. Data collected were subjected to analysis of variance (ANOVA) using Genstat statistical package (GenStat 6.1). Genetic variances for plant height, days to flowering, days to maturity, yield and yield components were determined by analysis of variance. From the angular leaf spot data, Chi-square was calculated to obtain Mendelian segregation ratios while heritability was calculated as the proportion of total genetic variance in a progeny. RESULTS Inheritance of angular leaf spot. All the plants of Lusaka Yellow were susceptible, while those of Mexico 54 were resistant. All the F1 plants showed resistant reactions to P. griseola (Table 3). The backcross to Lusaka Yellow showed 1:1 segregation ratio, while the backcross to Mexican 54 had all the progenies resistant. All Pembela plants were susceptible to the angular leaf spot while all Mexico 54 plants were resistant (Table 4). The F1 were all resistant, while the F2 segregated in the 3:1 ratio for resistance to susceptible. Backcrosses of the F1 to the susceptible parent produced progeny that showed a 1:1 resistant:susceptible ratio. However backcross progeny to the resistant parent were all resistant. Inheritance of quantitative.All the Lusaka yellow x Mexico 54 crosses and the parents showed significant differences in all the quantitative traits (days to flowering, days to maturity, plant height pod length and yield (Table 5). The degrees of dominance for plant height and number of pods per plant were 0.74 and 0.70 with additive genetic differences of 61%. Pod length showed a degree of dominance of 31%. The degree of dominance for number of seeds per pod was 0.28. Seed size of the F1 was better than the mid parent value by 6%. This trait showed an additive genetic difference of 65%, with a degree of dominance of .073. Grain Yield showed about 57% due additive genetic differences, while the degree of dominance was 0.45. Pembela x Mexico 54 crosses and parents were significantly different in days to maturity, plant height, pod and seed yield (Table 6). The crosses generally showed improved performance compared to parent 1. Pembela had slightly larger seeds compared to Mexico 54, but the seed sizes in the two parents were within the medium category of the CIAT classification (van Schoonhoven and Pastor-Corrales, 1987). An increase of 16% in seed size above the better parent and 21% above the mid parent value was observed in the F1. A heterosis of 11% in grain yield was observed above the better parent value. The crosses showed a higher heritability for number of pods per plant than grain yield (Table 7). DISCUSSION The F1 plants resulting from crosses between Lusaka yellow and Mexico 54 were all resistant (Table 3 and Table 4) indicating that the resistance was governed by single dominant gene. The F2 progenies segregated in a 3:1 ratio and the calculated Chi-square values gave a good fit for the ratio of 3 resistant to 1 susceptible. This type of segregation suggested that the resistance in Mexico 54 to angular leaf spot is governed by a single dominant gene as reported by Ferira et al. (2000) and Kimani et al. (2002). The BC1P1 showed 1:1 segregation ratio and the BC1P2 had all the progenies resistant. This agrees with the results by Namayanja (2003) and Mahuku et al. (2002) reported that the resistance in Mexico 54 was due to a single recessive gene. Mahuku et al (2003 and 2009) also reported three dominant genes (PhgG5686A, Phg G5686B,PhgG5686C ) in Mexico 54. Resistance to specific isolates of P.griseola has been reported to be simply inherited and molecular markers have been identified for some these resistance genes (Ferreira et al., 2000; Mahuku et al., 2004; Miklas et al., 2005). The non-significance of the 100 seed weight among the progenies of this cross indicated the closeness of the parents in terms of seed size. The dominance of genes from Mexico 54 governing days to flowering, maturity and plant height was observed in the progenies. Moreover, backcrosses to the donor parent had mean values above the mid parent, indicating the influence of the donor gene. The recombination of these two genotypes showed that it is possible to transfer resistance from the Mesoamerican to the Andean pool. The crosses showed a higher heritability for number of pods per plant compared to grain yield. This could indicate that selection for yield based on number of pods per plant would be less effective than based on the grain yield. Fernandez and Miller (1985), working with grain type cowpeas (Vigna unguiculata), also observed that pod number is the yield component most affected by the environment. The higher heritability observed in days to flowering, days to maturity, plant height, 100 seed weight and grain yield may indicates that the traits are useful in determining which family structure is best for maximising genetic gain over time (Holland et al., 2003). The lack of difference in segregation of the F2 generation for resistance to susceptible to Phaeoisariopsis griseola of the crosses Lusaka Yellow x Mexico 54 and Pembela x Mexico 54 from the expected 3:1 ratio (Table 3 and Table 4) indicates that the resistance to angular leaf spot in Mexico 54 is governed by a single dominant gene. The existence of significant positive heterosis in the yield and yield components in the crosses is encouraging as it indicates that gene combinations exist which can result in enhanced yield performance. The performance of the parents could be used to predict the performance of the progenies. The top yielders among the F1 had at least one of the parents with high grain yield per plot. This shows that for parents to enter a breeding programme they should be high yielding. High heritability values are important as they indicate that the selection of parents bearing particular measurements will produce progenies of the same phenotype (Holland et al., 2003). ACKNOWLEDGEMENT This research was supported by the Pan-Africa Bean Research Alliance (PABRA) and the University of Nairobi through grant No.751/2001. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11025t2.jpg] [cs11025t6.jpg] [cs11025t7.jpg] [cs11025t5.jpg] [cs11025t3.jpg] [cs11025t4.jpg] [cs11025t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}