 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 277-287 Resistance and inheritance of common bacterial blight in yellow beans. K.K. Muimui, P.M. Kimani1 And J.W. Muthomi1 Misamfu Regional Research Station, P. O. Box 410055, Kasama, Zambia
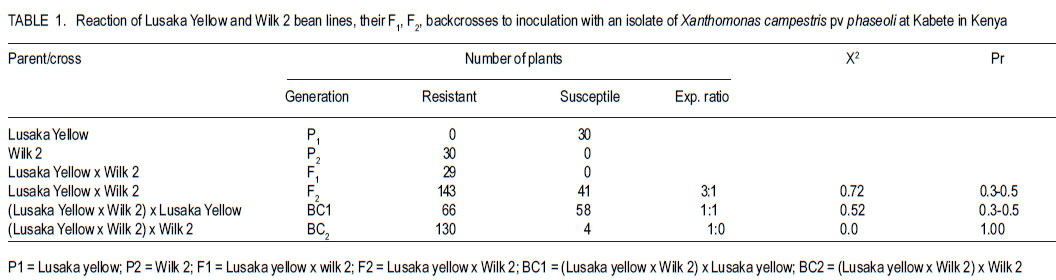
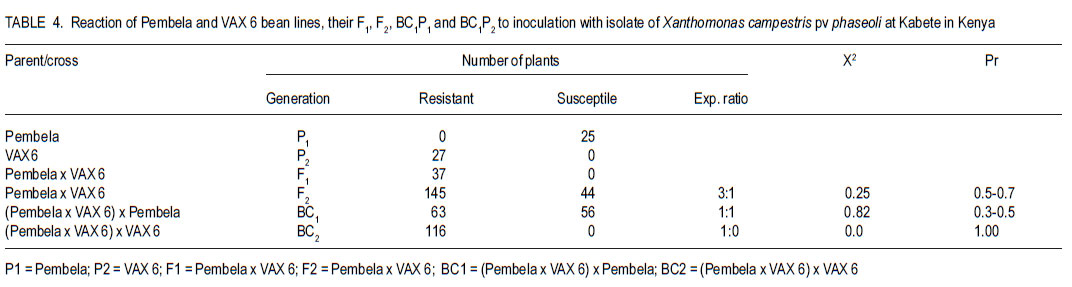
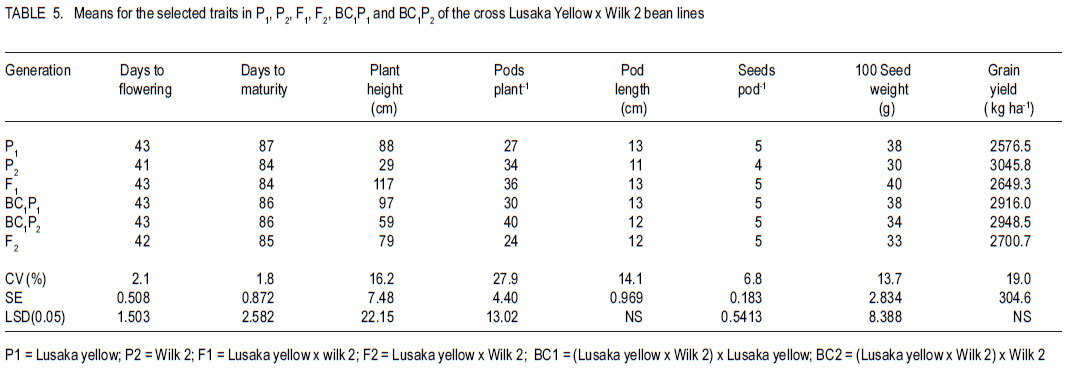
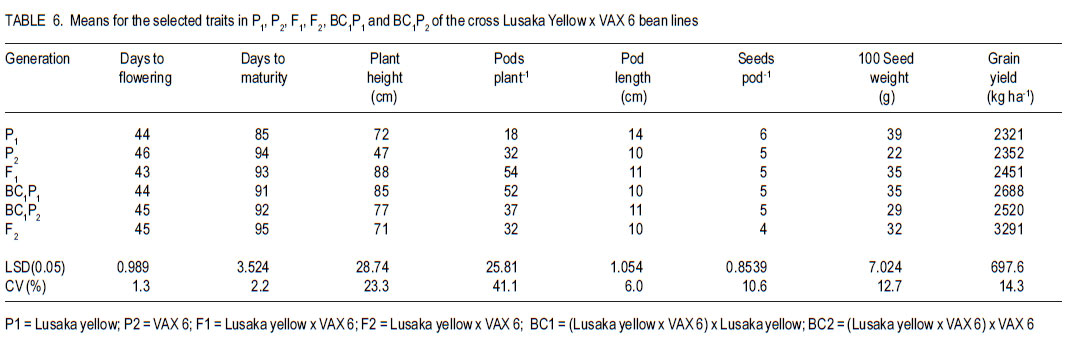
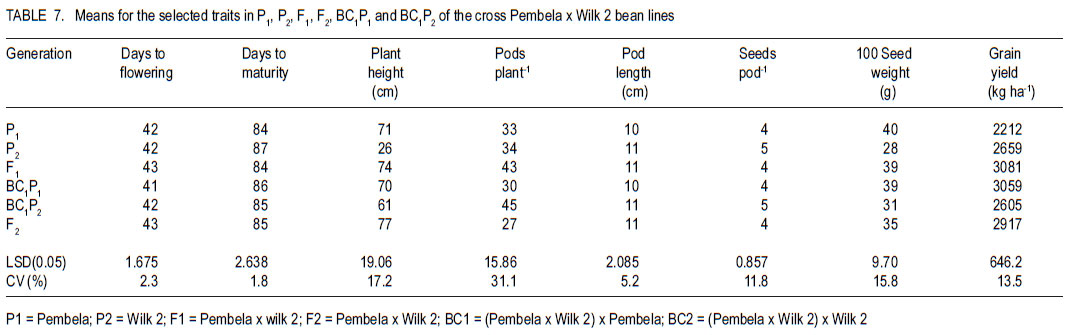
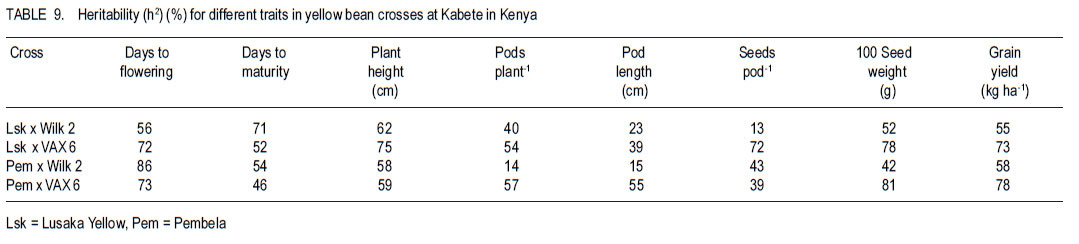
Code Number: cs11026 ABSTRACT Common bean (Phaseolus vulgaris L.) is the most important food legume among the pulses. It is a cheap source of protein, especially in Sub-Saharan Africa. However, bean production is constrained by bacterial diseases, of which common bacterial blight (Xanthomonas axonopodis p.v. phaseoli) is prevalent in Africa. The objective of this study was to transfer resistance to common bacterial blight and determine its inheritance in yellow beans. Sources of resistance were CIAT lines, Wilk 2 and VAX 6, which were crossed with susceptible Lusaka yellow and Pembla. The parents, F1, F2 and backcross progenies were inoculated with X. axonopodis and the resulting blight severity determined. Quantitative traits, including days to flowering, number of pods, and seed yield were also determined. The F1 and backcrosses to the resistant parents were all resistant, while the F2 and backcrosses to the susceptible parents segregated in 3:1 and 1:1 ratios, respectively. Additive genetic effects were observed in quantitative traits like days to flowering, plant height, days to maturity and yield. Therefore, resistance to common bacterial blight is controlled by a single dominant gene. The resistant parents Wilk 2 and VAX 6 could be used to improve bean varieties that are susceptible to common bacterial blight. Key Words: Phaseolus vulgaris, quantitative traits, Xanthomonas axonopodis RÉSUMÉ Le haricot commun (Phaseolus vulgaris L.) est une nourriture la plus importante parmi les légumineuses. C’est une source moins chère de protéines, spécialement en Afrique sub-Saharienne. Cependant, la production du haricot est handicapée par des maladies bactériennes, parmi lesquelles le flétrissement bactérien courant (Xanthomonas axonopodis p.v. phaseoli) qui est le plus prevalent en Afrique. L’objectif de cette étude était de transférer la résistance au flétrissement bactérien et de déterminer son hérédité dans les haricots jaunes. Les sources de résistance étaient lignées CIAT, Wilk 2 et VAX 6, qui étaient croisées avec le susceptible Lusaka jaune et Pembla. Les parents F1, F2 et les progénies de croisement en retour étaient inoculées avec X. axonopodis et le degré de sévérité du flétrissement déterminé. Des traits quantitatifs, incluant les jours à la floraison, le nombre de gousses et le rendement en grains étaient aussi déterminés. Les F1 et les produits de croisements en retour des parents résistants étaient tous résistants, alors que les F2 et les produits de croisements en retour des parents susceptibles étaient ségrégués dans les rapports 3:1 et 1:1, respectivement. Des effects génétiques additifs étaient observés dans les traits quantitatifs tels que les jours à la floraison, la hauteur des plants, les jours à la maturité et le rendement. Ainsi, la résistance au flétrissement bactérien est controllé par un seul gène dominant. Les parents résistants Wilk 2 et VAX 6 pourraient être utilisés pour améliorer les variétés de haricot susceptibles au flétrissement bactérien commun. Mots Clés: Phaseolus vulgaris, traits quantitatifs, Xanthomonas axonopodis INTRODUCTION Common bean (Phaseolus vulgaris L.) is the second most important source of dietary protein in Eastern and Southern Africa, where the consumption exceeds 50 kg person-1 year-1 (Pachico, 1993). In Africa, annual bean production is estimated at 4 million hectares (Wortmann et al., 1998). In addition to human nutrition, beans provide income for the small holder farmer through sale of surplus produce. Major seed classes grown include red mottled, large red kidney, small red, yellow, navy, purples, black and sugars (Wortmann et al., 1998). These classes are grown in different areas, depending on local preferences and market demand. Yellow beans are preferred because of their high price and short cooking time (Mulila-Mitti et. al., 1989). Widely grown yellow beans in Zambia include Pembela, Lusaka Yellow and Tabora. Lusaka Yellow is a local landrace variety grown in all bean growing regions of Zambia (Muimui, 2007). The variety is susceptible to angular leaf spot and common bacterial blight. Lusaka Yellow has an indeterminate upright type with medium seed size, which are yellow. It fetches premium prices in local markets in Zambia due to its colour and seed size. Pembela is a Zambian local variety which is also grown in most bean growing areas of the country (Muimui, 2007). Pembela is preferred for its colour and size, and cooks fast hence the name ‘Pembela’ meaning ‘wait’, because it cooks within a short time. It is susceptible to angular leaf spot and common bacterial blight. It is an indeterminate upright type with medium sized, yellow seeds. The main bean diseases in Zambia include angular leaf spot (Phaeoisariopsis griseola), anthracnose (Colletotrichum lindemuthianum), rust (Uromyces appendiculatus), common bacterial blight (Xanthomonas axonopodis p.v. phaseoli), and bean common mosaic and root rots. Common bacterial blight is prevalent in low to mid-altitude and warm areas (CIAT, 1996), causing yield losses of up to 75% (Allen et al., 1996; Wortmann et al., 1998). The effectiveness of cultural and chemical control methods is limited due to the high production costs. Therefore, the current focus is to develop varieties that have multiple-constraint resistance. Resistance to common bacterial blight has been reported in Phaseolus acutifolius (Park et al., 1998; Yu et al., 1998), P. coccineus and lines of P. vulgaris (Yu et al., 1998; Miklas et al., 1999). CIAT lines VAX 3, VAX 4, VAX 6, XAN 159 have also been reported to have good level of resistance to common bacterial blight (Singh and Muñoz, 1999). Moreover VAX 3, VAX 4 and VAX 6 possess levels of CBB resistance that are as high as those found in P. acutifolius accessions. Increased resistance can be developed by selecting for horizontal rather than vertical resistance (Garcia-Espinosa, 1997). Breeding yellow bean cultivars with resistance to major diseases would increase their productivity and improve returns for farmers. Therefore, the objective of this study was to determine the inheritance of resistance to common bacterial blight in segregating populations of yellow beans. MATERIALS AND METHODS Generation of experimental materials. The sources of resistance were CIAT lines Wilk 2 and VAX 6 from the Dry Bean Programme of the Agricultural Research Council of South Africa; while Lusaka yellow and Pembela were the susceptible parents. The susceptible parents were used as the females. Four crosses were made as follows: Lusaka Yellow X Wilk 2, Lusaka Yellow X VAX 6, Pembela X Wilk 2 and Pembela X VAX 6. Emasculation and pollination was done either early in the morning or evening as described by Tumwesigye (1988). The emasculated flowers were tagged and labelled with the pedigree of the cross. The resulting pods were harvested at physiological maturity, dried and threshed. The F1 seeds were planted to produce F2 and also backcrossed to the two parents. Pathogen culture and inoculation. Common bacterial blight pathogen (Xanthomonas axonopodis p.v phaseoli) was isolated from infected leaves and cultured on nutrient agar at 27 oC for 48 hours. The inoculum was harvested in sterile distilled water and adjusted to 109 colony forming units (CFUs) per millilitre by the plate count method. Plant leaves were artificially injured using a multiple needle puncher to create wounds for entry of the bacteria into the plant system. The inoculum was applied to the leaves of three-weeks old plants using a hand atomiser held at 10-15 centimetres from the leaf surface. A repeat inoculation was done at the start of flowering. Control plants were sprayed with sterile distilled water and all the plants were maintained in the inoculation chambers for the whole growing period and examined for disease symptom development. Experimental design. This study was conducted under greenhouse and field conditions at Kabete Field Station at the University of Nairobi in Kenya. In the greenhouse, three to four seeds of parents and their F1, F2 and backcross progenies were sown in plastic pots containing sterilised soil, sand and manure in a ratio of 3:1:1 v/v, respectively. Three to four seeds of each parent and F1 were planted in each pot and replicated three times. The BC1P1 and BC1P2 were planted into 14 pots per replication, while the F2 progenies were planted in 20 pots per replication. Seedlings were allowed to grow in inoculation chambers until maturity. The experimental design was completely randomised design with three replications. Sisal ropes were used to support the plants that showed some climbing tendency. In the field experiment, the parents (P1 and P), F1 , F2 and the backcross generations (BC1 P1 and BC1P2) were grown in a randomised complete block design, with three replications. The parents and F1 were sown in one row each per replication while four rows of the F2 generations and backcrosses were planted in each replication. The rows were 3 m long and 50 cm apart with an intrarow spacing of 15 cm. A total of 60 plants for each of the P1, P2 and F1 generations and 360 plants for the F2 , BC1 P1 and BC1 P2 generations were planted. Di-ammonium phosphate (DAP - 18% N and 46% P205) fertiliser was applied at planting time at the rate of 150 kg ha-1. Sticks of about 2 m long were used to support the climbing genotypes. Data collected included common bacterial blight severity, days to flowering and maturity, plant height, number of pods per plant, pod length, number of seeds per pod, number of seeds per plant, and 100 seed weight and seed yield. Disease assessment. Disease assessment started 15 days after inoculation or after the onset of symptom development. The assessment was based on a 1-9 disease severity scale (van Schoonhoven and Pastor-Corrales, 1987). Two assessments were done in the greenhouse and each plant was assessed by scoring three trifoliate leaves starting from the base. A mean score was calculated for each plant to determine the level of reaction to the pathogen. Data analysis. GenStat statistical package (Genstat 6.1) was used to analyse the data. Chi-square method was used to test significance of observed segregation ratios. Analysis of variance (ANOVA) was used to estimate genetic variances for quantitative traits. The disease severity data were subjected to qualitative genetic analysis using a chi-square to compare the Mendelian segregation of observed to hypothetical ratios. Characterisation of the genetic resistance was determined after analysing the segregating ratios obtained from the disease phenotypic reactions of the F2 populations and backcross generation. Heritability, which is the proportion of the total variation in a progeny that has genetic basis, was also calculated. RESULTS Inheritance of qualitative traits. All Lusaka Yellow plants showed susceptible reactions to common bacterial blight, while all the plants of Wilk 2 were resistant (Table 1). All the F1 plants were resistant to common bacterial blight, while the F2 segregated in a 3:1 ratio for resistant to susceptible. The results in BC1P1 were 50% resistant and 50% susceptible and all the plants in the BC1P2 were resistant. In the crosses between Lusaka Yellow and VAX 6, all the VAX 6 plants showed resistance reaction to common bacterial blight, but the all the F1 plants were resistant (Table 2). The F2 plants segregated in a 3:1 ratio for resistant to susceptible; while the backcross segregated in the ratio of 1:1 and were all resistant. Crosses between Pembela and Wilk 2 showed that all the Pembela plants were susceptible to the bacterial blight, while the Wilk 2 plants were all resistant (Table 3). All the F1 plants were resistant and the F2 plants segregated in the 3:1 ratio for resistant and susceptible. Half of the backcross to the susceptible parent was resistant with the other half showed susceptible, representing a 1:1 ratio. The crosses between Pembela and VAX 6 showed that all Pembela plants were susceptible, while all the VAX 6 plants were resistant (Table 4). In F1, all the plants showed resistant reaction, while the F2 plants segregated in the 3:1 ratio for resistant to susceptible. Back cross to the susceptible parent segregated in the 1:1 ratio and the plant resulting from the backcross to the resistant parent showed resistant reaction. Inheritance to quantitative traits. In the crosses between Lusaka Yellow x Wilk 2, dominance of days to flowering accounted for about 86%, yet 71% of the variability in duration to maturity was due to additive genetic effects (Table 5). The number of pods per plant for the F1 was better than the better parent by 6%. Lusaka Yellow had more seeds per pod and larger seeds than Wilk 2. The F1 progeny showed heterosis for 100-seed mass. Seed weight in the F1 was improved by 17% above the mid parent value. Wilk 2 yielded more than Lusaka Yellow. However, yield of the F1 progeny showed no heterosis for grain yield. The degree of dominance for this trait was 0.36 and the effects due to additive genetic differences accounted for about 55%. The additive genetic difference for plant height in crosses between Lusaka Yellow x VAX 6 accounted for 75% with a degree of dominance of 0.50 (Table 6). The degree of dominance for the number of pods per plant was 0.72, while the additive genetic differences for pod length accounted for 39%. The yield for the F1 obtained from this cross was better than both parents, with a mean yield of 2451 kg ha-1. The degree of dominance for this trait was 0.43. The degree of dominance for plant height and number of pods per plant in crosses between Pembela and Wilk 2 was 0.77 and 0.49, respectively, while that for pod length was 0.74 (Table 7). The additive genetic difference for the number of seeds per pod was about 0.43, while the degree of dominance for this trait was 0.27. Heterosis was observed in this cross with better yield for F1 than the parents. Heterosis of 27% was observed above the mid parent value and 16% above the better parent value. Effects due to additive genetic differences were 58%, while the degree of dominance was 0.44. The degree of dominance for plant height in Pembela x VAX cross was about 0.65 and the number of pods per plant for the F1 hybrid was above the better parent by 78% and above the mid parent value by 95% (Table 8). The number of pods per plant had a dominance of 0.70, while that for pod length was 0.36, of which 55% was due to additive genetic differences. In the case of the number of seeds per pod, the degree of dominance was 0.61 and an additive genetic difference of 39%. A heterosis of 8% above the mid-parent value was observed with a degree of dominance of 0.56 (Table 9). DISCUSSION As expected, all Wilk 2 and VAX 6 plants were resistant to common bacterial blight, while all of Lusaka Yellow and Pembela were susceptible (Table 1 , Table 2, Table 3, Table 4). In the F1, all plants were resistant, while in F2 they segregated in the 3:1 for resistant to susceptible indicating that resistance was controlled by dominant genes. This is in line with the findings of Singh and Muñoz (1999), which showed that VAX 6 had high levels of resistance to common bacterial blight. The segregation of the F2 generation for resistance to susceptible to X. axonopodis in the crosses Lusaka Yellow x Wilk 2, Lusaka Yellow x VAX 6, Pembela x Wilk 2 and Pembela x VAX 6 did not differ from the expected 3:1 ratio indicating that the resistance to common bacterial blight in Wilk 2 and VAX 6 could be governed by a single dominant gene. The dominant nature of inheritance in the donor lines used in this study could make transferring common bacterial blight resistance from the cultivars Wilk 2 and VAX 6 relatively easy. The BC1P1 segregated in the ratio of 1:1 and all the BC1P2 progenies were resistant (Table 1 , Table 2, Table 3, Table 4). This is an indication of a possible dominant single gene for resistance to X. axonopodis in Wilk 2 and VAX 6. This is in line with the results of Singh and Muñoz (1999), which showed that Wilk 2 and VAX 6 have high levels of resistance to common bacterial blight. Furthermore, Fourie (2002) reported high levels of resistance in Wilk 2 and VAX 6 when they were used in a bean breeding programme in South Africa. Therefore, Wilk 2 and VAX 6 would be valuable sources of of resistance to common bacterial blight and the resistance genes can be incorporated to susceptible varieties by backcrossing to the resistant lines. The nature of inheritance greatly depends on the genotype used as the susceptible parent among other factors (Pastor–Corrales et al., 1998). In addition, Silva et al. (1989) reported that inheritance of resistance to common bacterial blight in trifoliate leaf and plant canopy was controlled by a single major gene. However, Musana et al. (1993), who worked on 10 crosses between resistant and susceptible bean genotypes, found that resistance to common blight was controlled by two or more genes. Therefore, resistance to common bacterial blight is different depending on the source of resistance and may be determined by both major and minor genes (McClory, 1985) . There are several other reports of resistance transferred from P. acutifolius to P. vulgaris (Thomas and Waines, 1984; Park et al., 1998; Yu et al., 1998). Resistance in P. acutifolius is controlled by one or two dominant genes and is related to the hypersensitive response (Zapata, 1998; Urrea et al., 1999). In addition, crosses between P. coccineus and P. vulgaris also showed resistance to X. axonopodis p.v.phaseoli (Zapata et al., 1985; Yu et al., 1998). A number of P. vulgaris lines with varying levels of resistance to X. axonopodis p.v. phaseoli have been registered (Miklas et al., 1999). Partial resistance to X. axonopodis in P. vulgaris has been linked to delayed flowering under long photoperiods (Goodwin et al., 1995). There are also several other reports of resistance transferred from P. acutifolius to P. vulgaris (Thomas and Waines, 1984; Park et al., 1998; Yu et al., 1998). In this study, duration to maturity was controlled by additive genes (Table 5, Table 6, Table 7). Lusaka Yellow had a higher number of seeds per pod and larger seeds than Wilk 2, and the F1 progeny showed heterosis for 100-seed mass (Table 7). The F2 progenies had a wider range of seed sizes and the 100-seed mass varied from 16 to 50 g. This could be attributed to segregation of genes controlling seed size (Upadhyaya et al., 2006). Genetic analyses showed that seed size was largely controlled by additive genes (Table 8). More than 52% of the variability was due to additive gene action. This is in agreement with Cho et al. (2002) who identified quantitative trait loci (QTL) that accounted for 52% of total phenotypic variation for seed size in chickpea. The existence of significant positive heterosis in yield and yield components in the crosses indicates that gene combinations exist, which can result in enhanced yield performance. The gene for indeterminate was dominant to the determinate as most of the crosses that were made between the two types resulted in progenies way above the mid parent value. High heritability values are important, as they indicate that the selection of parents bearing particular measurements will produce progenies of the same phenotype. From this study, it possible to improve local bean landraces such as Pembela and Lusaka Yellow. ACKNOWLEDGEMENT This research was supported by the Pan-Africa Bean Research Alliance (PABRA) and the University of Nairobi through Grant No.751/2001. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11026t7.jpg] [cs11026t5.jpg] [cs11026t9.jpg] [cs11026t3.jpg] [cs11026t8.jpg] [cs11026t6.jpg] [cs11026t1.jpg] [cs11026t4.jpg] [cs11026t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}