 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 289-302 Multiple disease resistance in snap bean genotypes in Kenya. S.W. Wahome, P. M. Kimani, J.W. Muthomi, R.D. Narla and R. Buruchara1 Department of Plant Science and Crop Protection, University of Nairobi, P. O. Box 29053-00625
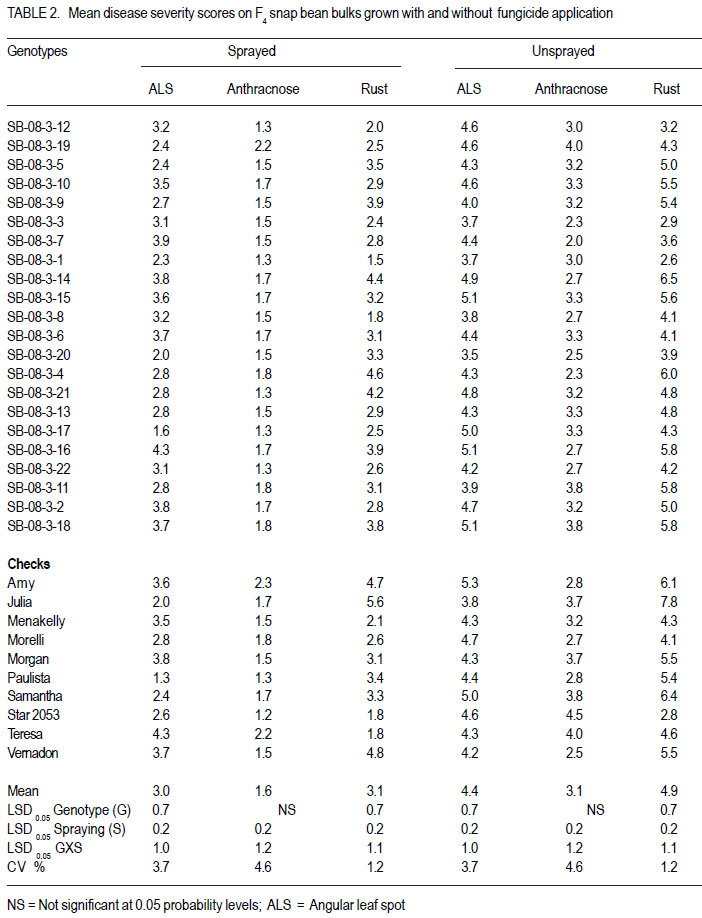
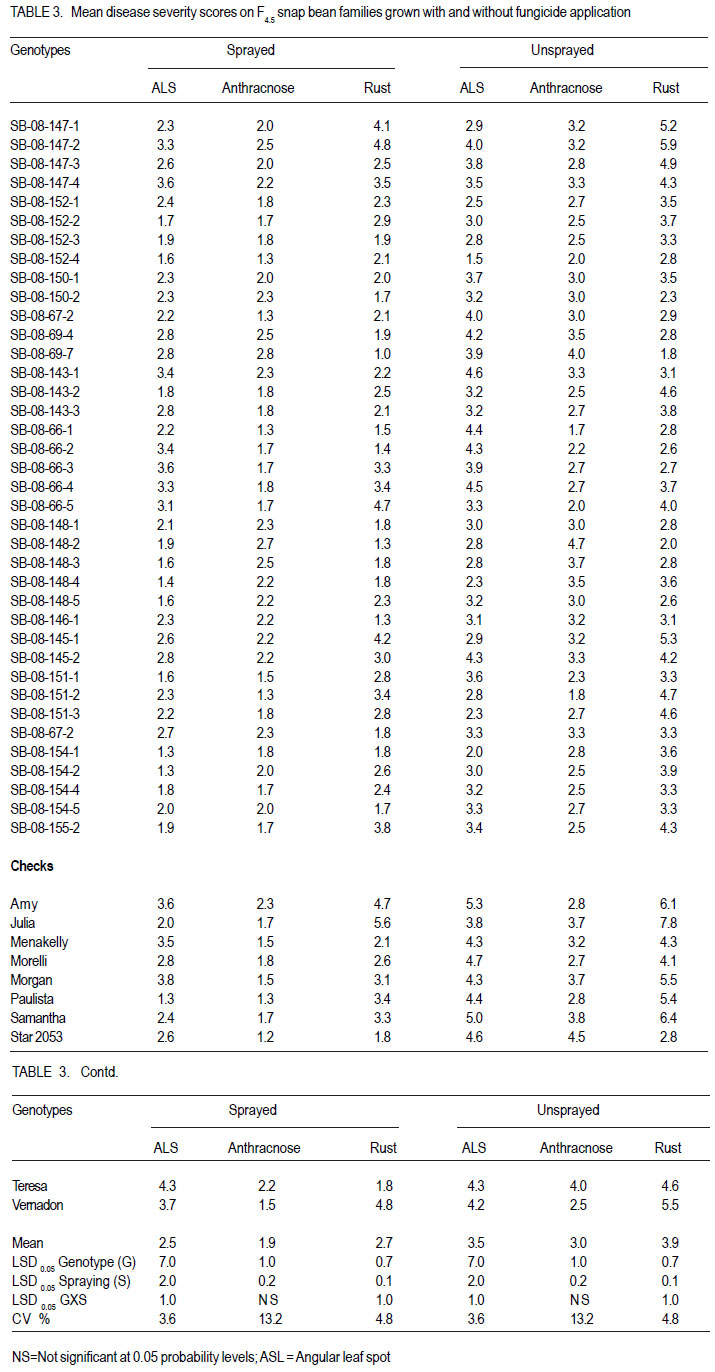
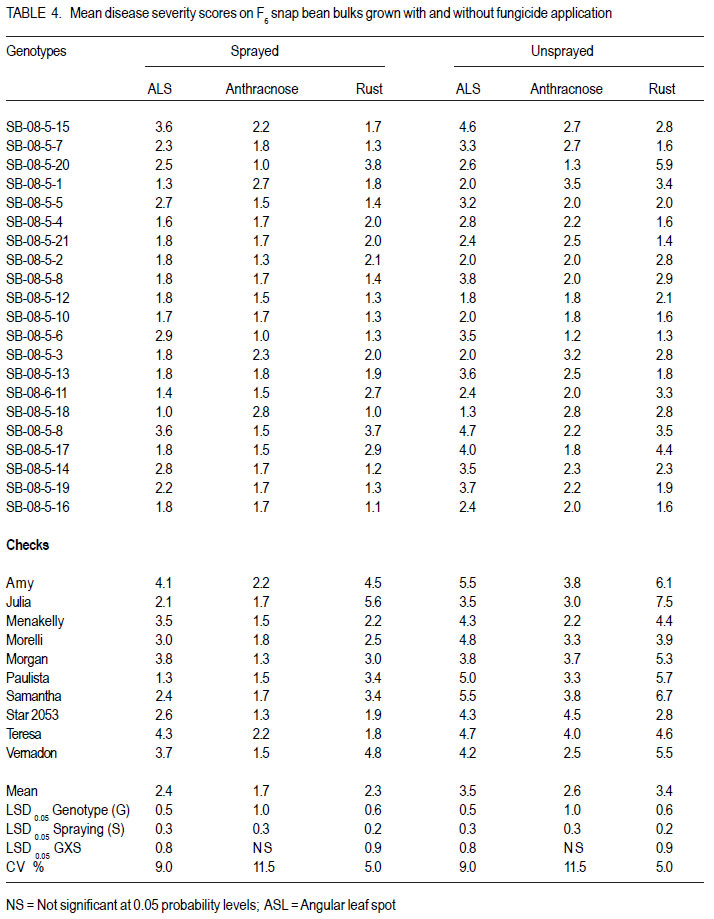
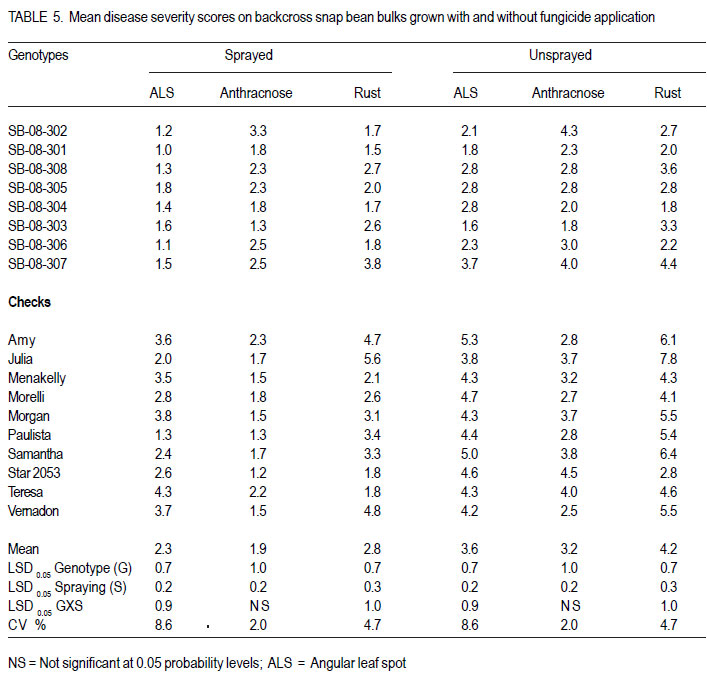
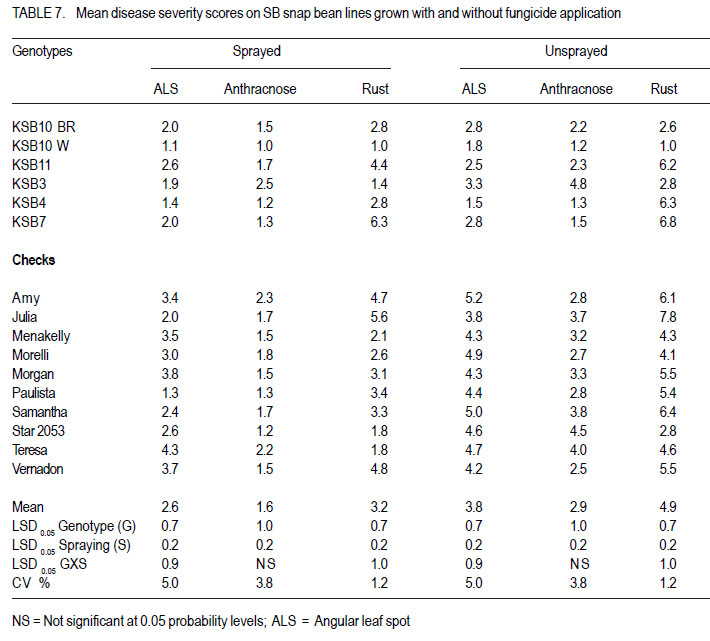
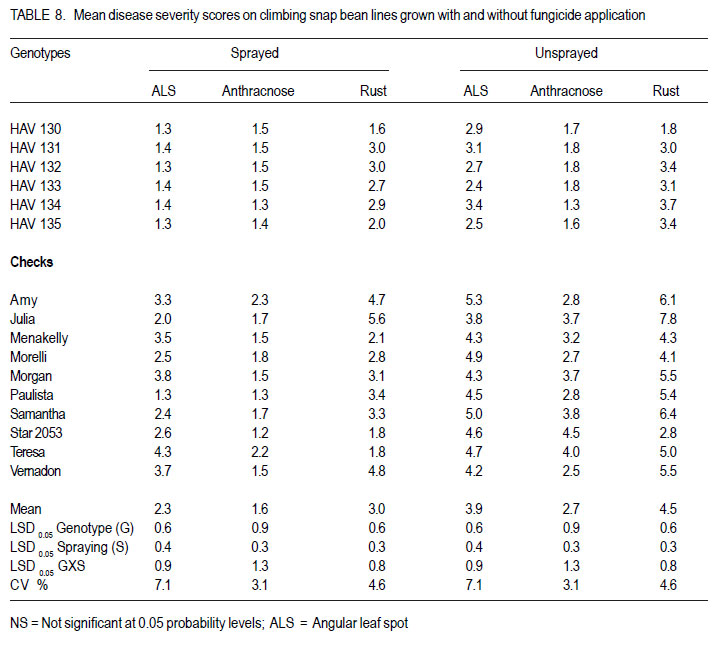
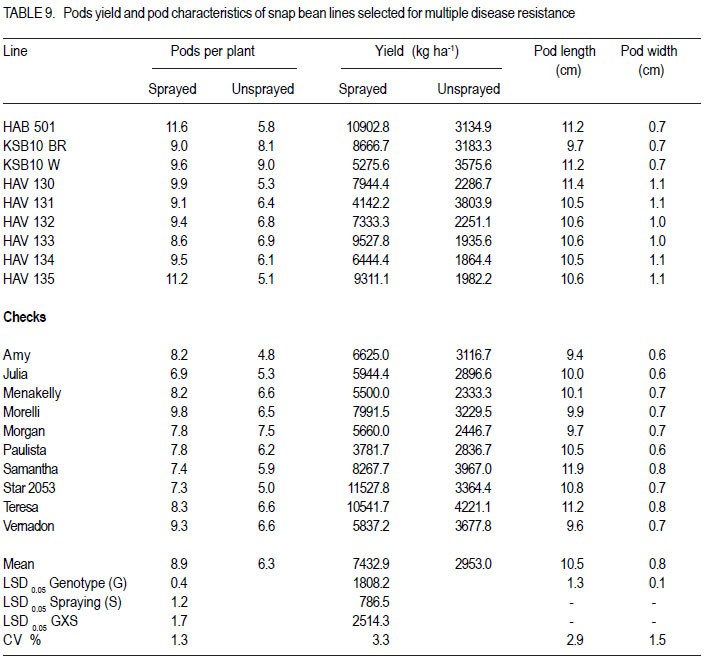
Nairobi, Kenya Code Number: cs11027 ABSTRACT Snap bean (Phaseolus vulgaris L.) is an important export vegetable crop, produced mainly by small to medium scale farmers under various disease constraints. Disease resistant varieties can reduce reliance on fungicides, and enhance the capacities of smallholder farmers to meet the stringent European export requirements for allowable fungicide residues. This study was carried out to identify snap bean lines with multiple disease resistance to angular leaf spot (Phaeoisareopsis griseola), anthracnose (Collectotrichum lindemuthianum) and rust (Uromyces appendiculatus). Seven groups of snap bean populations of different generations, and 45 bush snap bean lines, including local checks, were evaluated for resistance to the three diseases at two locations in Kenya. The disease with the highest severity was rust, followed by angular leaf spot. Among the advanced lines, two bush lines (KSB 10 W and KSB 10 BR), and one climbing line (HAV 130) had consistent multiple resistance to angular leaf spot, anthracnose and rust at both locations. Nine lines and 674 single plants were selected from populations showing multiple disease resistance. Resistance in selected lines reduced angular leaf spot, anthracnose and rust severity by 17, 16 and 36%, respectively. The multiple disease resistant lines were not the highest yielders but had the highest number of pods per plant. Climbing snap bean lines had thick pods that could reduce pod quality. Key Words: Angular leaf spot, anthracnose, rust RÉSUMÉ Le haricot vert (Phaseolus vulgaris L.) est une importante légume d’exportation, produite principalement par de petits fermiers sous différentes contraintes de maladies. Des variétés résistantes aux maladies peuvent réduire l’utilisation de fongicides, et promouvoir la capacité des petits fermiers à satisfaire les exigencies rigoureuses des exportations européennes en résidus acceptables de fongicides. Cette étude était ménée afin d’identifier des lignées de haricots verts avec multiple résistance à la tâche angulaire des feuilles (Phaeoisareopsis griseola), à l’anthracnose (Collectotrichum lindemuthianum) et à la rouille (Uromyces appendiculatus). Sept groupes de populations de différentes générations, et 45 lignées de haricots verts nains incluant les témoins locaux, étaient évalués pour résistance aux trois maladies dans deux localisations au Kenya. La maladie avec une sévérité la plus élevée était la roulle, suivie par la tâche angulaire des feuilles. Parmi les lignées avancées, deux lignées naines (KSB 10 W et KSB 10 BR), et une volubile (HAV 130) avaient une resistance multiple consistante à la tâche angulaire des feuilles, à l’anthracnose et à la rouille dans toutes les deux localisations. Neuf lignées et 674 plantes simples sélectionnnées des populations avaient montré une résistance multiple aux maladies. La résistance dans les lignées sélectionnées avaient réduit la séverité de la tâche angulaire des feuilles, de l’anthracnose et de la rouille de 17, 16 et 36%, respectivement. Les lignées de résistance multiple n’avaient pas des rendements plus élevés mais avaient un nombre plus élevé de gousses par plante. Les lignées de haricots verts volubiles avaient des gousses épaisses susceptibles à réduire la qualité de gousses. Mots Clés: Tâche angulaire des feuilles, anthracnose, rouille INTRODUCTION Snap bean (Phaseolus vulgaris L.) is an important export vegetable crop in Africa. More than 90% of the crop produced in Eastern Africa is exported to regional and international markets. Production is mainly by small to medium scale farmers and the enterprise creates on-farm employment opportunities for the rural community. More than one million people benefit from the snap bean sub-sector in Kenya (CIAT, 2006). However, access to the European Union markets is becoming increasingly difficult as a result of strict safety and quality standards (Monda et al., 2003). Snap bean farmers face several constraints, which include pests, diseases and stringent quality requirements (Monda et al., 2003). Rust (Uromyces appendiculatus), angular leaf spot (Phaeoisareopsis griseola) and anthracnose (Collectotrichum lindemuthianum) are the most important and widely distributed diseases of snap beans in eastern Africa. Angular leaf spot causes severe and premature defoliation resulting in shriveled pods, shrunken seeds and yield losses of up to 80% (Stenglein et al., 2003). Bean rust is endemic and severe in eastern and southern Africa and causes yield and quality reductions ranging from 18 to 100% in humid and tropical areas (Kimani, 2002; Monda et al., 2003). Bean anthracnose is transmitted from one season to another through infected seed and when infection occurs early in growth cycle of susceptible cultivars, yield loss can be up to 100% (Fernandez et al., 2000). Smallholder snap bean farmers mainly rely on fungicides and insecticides to reduce production and post-harvest losses associated with diseases (Wasonga et al., 2010). However, the use of chemicals makes the produce less marketable due to the maximum chemical residue level requirements set by the European markets (Kimani, 2002). Continued use of chemicals also leads to emergence of disease resistant pathogen races, increased production costs and negative effect on the environment and human health (Burkett-Cadena et al., 2008). Cultural practices such as crop rotation, intercropping,elimination of plant debris, adjustment of planting dates, use of compost, and blending heterogeneous cultivars can reduce diseases severity (Deeksha et al., 2009). However, the use of host plant resistance is by far the most economic and environmentally sustainable method for controlling bean diseases. Therefore, the development of cultivars with improved resistance to biotic and a biotic stresses has been the goal in bean breeding (Milkas et al., 2002; Sharma et al., 2007). This entails development of snap bean varieties with multiple resistances to rust, anthracnose and angular leaf spot diseases. The objective of this study was to select snap bean populations and advanced snap bean lines for multiple disease resistance to rust, angular leaf spot and anthracnose, pod quality and yield. MATERIALS AND METHODS Experimental site. Field experiments were conducted at the Kenya Agricultural Research Institute in Thika (KARI-Thika) and on farmer’s field in Mwea in Kenya. KARI-Thika is located in co-ordinates 00 59’ South and 370 04’ East at an elevation of 1548 m above sea level. It experiences bimodal pattern of rainfall with an annual mean of 1000 mm. Long rains occur between March and May while short rains occur between October and December with a mean of 142 mm and 116 mm, respectively. The mean annual maximum and minimum temperatures are 25.1 and 13.7 °C, respectively (Ndegwa et al., 2009). Mwea is located at 37o 20’ East and at an elevation of about 1159 m above sea level. It experiences bimodal rainfall with an annual mean of 1037 mm with long rains occurring between March and May while short rains are between October and December with a mean of 71 mm to 50 mm, respectively. The mean annual maximum and minimum temperatures are 27.8 °C and 15.6 °C respectively (Ndungu et al., 2004). Plant materials and experimental design. Climbing and bush snap beans lines and advanced segregating populations developed from BelDakMi, L227, Beltigrade RR2, Awash 1, G2333, BelMiNeb and Roba-1 and nine susceptible commercial varieties (Amy, Paulista, Morelli, Morgan, Julia, Foskelly, Teresa, Vernandon, Kutuless and Alexandria) were evaluated for resistance to angular leaf spot, anthracnose and rust. The F1 derived from the crosses were advanced to F4, F5 and F6 generations by the bulk population method and the three generations were subjected to evaluation. The experiment was laid out in a randomised complete block design with a split plot arrangement. The experiment was replicated three times at each site and for two seasons. Fungicide application was the main plots, while the snap bean genotypes were the subplots. The sprayed plots were treated every 10 to 15 days with Thiovit® 80 WG (sulfur) and Ortiva® (azoxystrobin) at the recommended rate of 1 ml L-1 of water. The snap bean populations and advanced lines were sown in two-row plots per replication. Each row was 3 m long and 50 cm apart. The distance between plants was 15 cm leading to a total of 42 plants for each genotype per plot. A 2- m long string or sticks was used to support the climbing genotypes. The first hand weeding was done two weeks after emergence and the second one just before flowering. Insect pests were controlled by alternate application of Dimethoate® (Deltamethrin) and Karate® (Lambda cyhalothrin) at the rate of 1.5ml L-1 every week. Pathogen isolation and plant inoculation. Colletotrichum lindemuthianum and Phaeoisariopsis griseola were cultured on potato dextrose agar and V8 juice agar, respectively. The inocula were harvested in sterile distilled water and spore concentration adjusted to 2 x 106 spores ml-1. Uromyces appendiculatus was multiplied and maintained in the field on susceptible snap bean varieties Teresa, Samantha, Amy, Paulista and Julia. Inoculation was by spraying the spore suspension on the leaves of 15-day old plants using a household atomiser. Disease severity assessment. Disease assessment was initiated 15 days after inoculation and was based on a 1-9 severity scale according to Van Schoonhoven and Pastor-Corrales (1987) (Table 1). Five plants were randomly sampled per plot for each genotype and assessed for angular leaf spot, rust and anthracnose severity by scoring three trifoliate leaves sampled at bottom, middle and top of each plant. Disease scores were recorded every two weeks until maturity. Mean disease scores were calculated for each plant and used to determine the level of reaction to the pathogen. Yield and yield components. Thirty plants per plot were randomly selected and mature pods were harvested and graded into three standard categories defined by width of the pod as extra fine (6 mm), fine (6-8 mm) and bobby (>8 mm) and length of the pods above 10 cm (HCDA, 2009). Harvesting for both bush and climbing snap beans was done three times per week at two day interval for 4 to five weeks. The fresh weight of each grade was determined at each harvest and the cumulative yield for the six weeks calculated at the end of the harvest period. For each pod grade, the number of pods per plant, pod length and pod width was also determined. Pod length and width was determined using a special ruler manufactured by Royal Sluis with holes of diameter 6, 8 and 12 mm for extra fine, fine and bobby pods, respectively. Data analysis. Quantitative data collected from the experiment were subjected to combined analysis of variance (ANOVA) using the PROC ANOVA procedure of GenStat (Lawes Agricultural Trust Rothamsted Experimental Station 2006, Version 9). Genotypes and fungicide treatments were treated as fixed effects. Location was considered to be a random effect. Differences among the genotypic means, and between fungicide treatments were compared using the Fisher’s protected LSD test at 5% probability level. RESULTS There were significant (P<0.05) differences among the F4, F6, and backcross populations, and among HAB lines, SB lines and climbing snap lines in angular leaf spot, rust and anthracnose severity (Table 2, Table 3, Table 4). However, F4.5 families had significant difference in angular leaf spot and rust severity only (Table 3). Bean rust had the highest mean severity scores, followed by angular leaf spot for most of the populations. In contrast, angular leaf spot was the most severe among the HAB lines followed by rust. Spraying with fungicides significantly reduced the severity of all the three diseases (Table 4) and the interaction between genotype and spraying was also significant. The snap bean varieties and segregating populations showed differences in reaction to anthracnose, angular leaf spot and rust. Lines showing high resistance to anthracnose were segregating populations like SB-08-3-5, SB-08-52 and SB-08-5-7. Varieties Morgan, Paulista, Star 2053, Teresa and Menakelly; and populations SB08-3-19, SB-08-148-3, SB-08-69-7 and SB-08-302 showed intermediate resistance to anthracnose (Table 5). Variety Julia and most segregating populations, bush and climbing lines, showed high resistance to angular leaf spot while most of the parent lines had intermediate resistance (Table 6). Variety Star 2053 was the only parental line with high levels of resistance to rust but many segregating populations, advanced bush and climbing lines showed high levels of resistance to rust. Variety Julia was the most susceptible with a score of up to 7.8. None of the parental lines had multiple resistance to all the three diseases but several segregating populations such as SB-08-3-3, SB-08-3-1, SB-08-3-1, SB-083-20, SB-08-302, SB-08-307 showed combined resistance to anthracnose, angular leaf spot and rust (Table 7 and Table 8). A few advanced lines such as HAB 501, SB 10 BR and SB 10 W showed multiple disease resistance. There were significant differences among the genotypes selected for multiple disease resistance in the number of pods per plant, pod yield, pod length and pod width (Table 9). Spraying significantly increased the number of pods per plant and pod yield. The interaction of genotype and fungicide application had a significant effect on the number of pods per plant and pod yield. Among the advanced lines, SB 10 W and SB 10 BR had the highest number of pods per plant while HAV 131 and SB 10 W had the highest pod yield. Pod length varied from 9.4 cm to 11.9 cm among the parental lines. Pod length varied from 9.7 cm to 11.4 cm among the advanced lines and segregating populations. Among the parental lines, Samantha had the longest pods while HAV 130 had the longest pods among the selected lines. Climbing lines had the highest pod width compared to the bush lines and parental varieties (Table 9). DISCUSSION Appearance of significant differences for the diseases severity among the populations shows that variability for resistance existed among the genotypes for rust and angular leaf spot and anthracnose. This indicates a possibility of obtaining genotypes with genes for resistance from their donor parents and confirms presence genes for resistance in the donor parents used in this study as reported by Markell et al., (2009) and Pastor-Corrales et al., (2010). Based on the mean disease severity scores, 674 single plants with multiple disease resistance were selected from the populations and families. However, susceptible plants would appear in the next generation from some of the selected single plants indicating that some were still segregating. Advanced lines HAB 501, SB 10 W and SB 10 BR and climbing lines HAV130, HAV131, HAV132, HAV133 HAV134 and HAV135 showed multiple commercial bush varieties. Such genotypes that resistance to rust, angular leaf spot and possess multiple trait resistance are useful since anthracnose. These nine lines reduced angular farmers are exposed to various diseases that leaf spot, anthracnose and rust severity by 17, 16 contribute to yield loss (Pascal et al., 2010). The and 36%, respectively, compared to the results indicate the advanced lines possess multiple disease resistance genes that could be useful in breeding programmes as has been reported by Fininsa and Tefera (2006) who found resistance to anthracnose, angular leaf spot and common bacterial blight. However, the parental varieties used as checks in the current study did not exhibit multiple resistance to the three diseases but Teresa showed intermediate resistance to rust and had highest total pod yield. This confirms the ur-5 genes for resistance to rust in Teresa as reported by Pastor-Corrales, (2006) and this gene is somehow effective in Mwea and Thika. Rust followed by angular leaf spot had the highest disease severity which could be explained by the high virulence and variability in the bean rust pathogen. Over 300 races or pathotypes of rust are recognised indicating the broad variability of the rust fungus (Araya et al. 2004). Under favourable environment found at the study sites, the pathogen is capable of rapidily overcoming newly deployed resistance. The high pathogenic variability of the rust fungus has been reported to make most of snap bean cultivars grown in Eastern and Southern Africa to be susceptible by Hillocks et al. (2006) and Monda et al., (2003). Therefore, development of snap bean varieties with several genes of resistance to rust is therefore important in order to control the disease. While other genotypes were highly susceptible to rust, HAB lines were more susceptible to angular leaf spot (Table 6). This suggests presence of rust resistance genes in these genotypes, but lacked genes for resistance to angular leaf spot. Resistance to Andean and Mesoamerican races of angular leaf spot has also been reported by Mahuku et al., (2003). Resistance to angular leaf spot is reported to be controlled by a single dominant gene (Fereira et al., 2000; Kimani et al., 2002) while Mahuku et al. (2009) reported that the resistance is controlled by three dominant genes PhgG5686A ,Phg G5686B and Phg G5686C. The results revealed that application of fungicide is an effective way of controlling the diseases, saving about half of the yield loss due to diseases. However, fungicides are expensive and are not environmentally friendly (Fontem et al., 2007). Therefore an integrated approach of growing resistant varieties with minimal use of fungicide would reduce yield loss caused by the diseases and also relieve farmers the high cost of applying fungicide. The genotypes experienced less rainfall and higher temperatures at both locations and this influenced the development of diseases. For example, Anthracnose is favoured by high temperatures of 26 oC, relative humidity of 85% and combined with frequent heavy rainfall (Mohammed and Somsiri, 2007). Lines SB 10 W and SB 10 BR had high number of pods per plant and this could be attributed to the fact that the two lines are resistant to anthracnose, angular leaf spot and rust while Amy had the least number of pods per plant since it was highly susceptible the three diseases (Table 9). Pod length of the commercial varieties was slightly shorter than the optimum size. This indicates that conditions in the test sites such as long period of moisture stress, low soil fertility and modest fertiliser application rates may have influenced expression of this trait. Only genotypes with a minimum pod length of 10 cm are suitable for export market (Muchui et al., 2001; HCDA, 2009). Among the selected lines, only SB10 BR had a pod length below 10 cm. Climbing lines had the highest pod diameter compared to the bush lines when they were harvested at regular intervals. This indicates that climbing snap bean could be harvested more often than the bush to avoid overgrown pod or they should be crossed with bush snap bean to incorporate thin pod characteristic. Bush lines had a pod width comparable to that of commercial varieties. Variety Teresa yielded well even under disease stress, indicating that it has some tolerance to the diseases infection. The results of the study indicated that the parental snap bean lines were successful in transferring genes of resistance. However, the selected climbing lines had thick pods that could limit them producing high proportion extra fine export quality pods. ACKNOWLEDGEMENT Funding for this study was from CIAT/Pan-Africa Bean Research Alliance (PABRA), while facilities and support provided by the University of Nairobi Bean Program and the Kenya Agricultural Research Institute (KARI-Thika) are gratefully acknowledged. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11027t6.jpg] [cs11027t9.jpg] [cs11027t1.jpg] [cs11027t5.jpg] [cs11027t3.jpg] [cs11027t7.jpg] [cs11027t2.jpg] [cs11027t8.jpg] [cs11027t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}