 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 325-333 Inheritance of halo blight resistance in common bean B.Y.E. Chataika, J.M. Bokosi, R.M. Chirwa1 and M.B. Kwapata Bunda College of Agriculture, P. O. Box 219, Lilongwe, Malawi
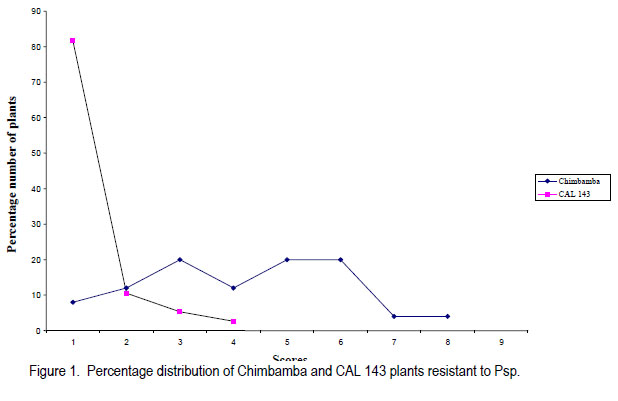
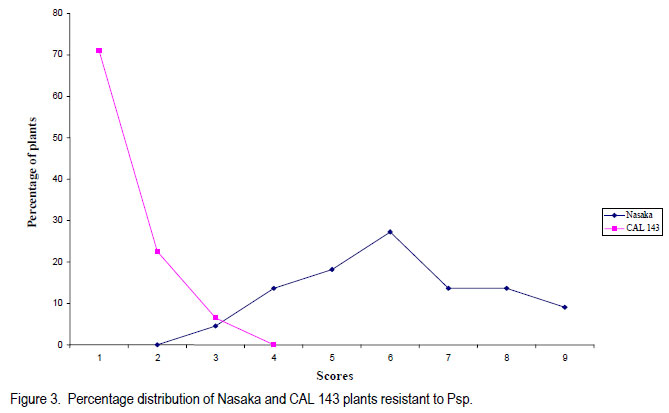
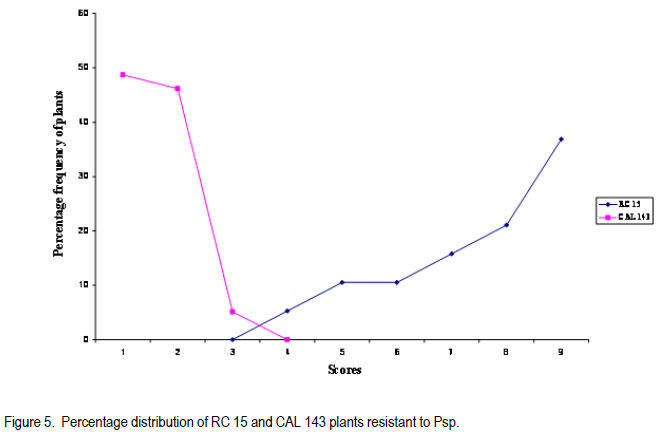
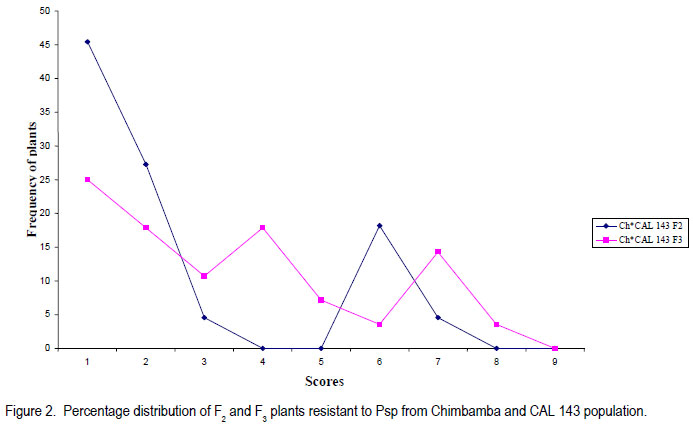
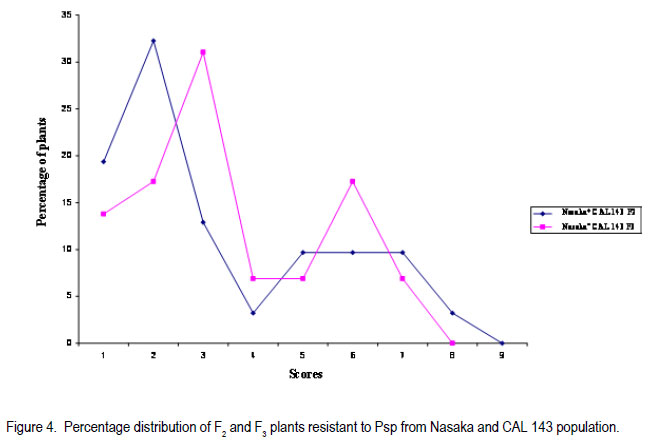
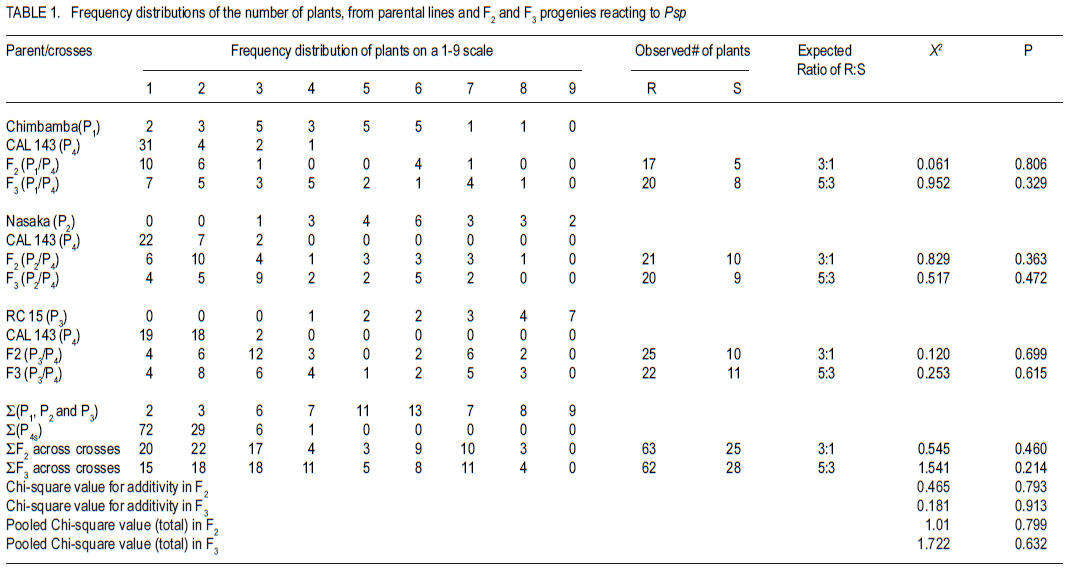
Code Number: cs11030 ABSTRACT Halo blight caused by (Pseudomonas syringe pv. phaseolicola (Burkh) (Psp)) is an important disease of common bean (Phaseolus vulgaris L.) world-wide. Several races of the Psp exist and likewise some sources of resistance in common bean have been identified. CAL 143, is a CIAT-bred common bean line, which was released as a variety in Malawi and several other countries in southern Africa. It possesses good resistance to Psp. However, the mode of inheritance for resistance to Psp in CAL 143, which is essential for devising a proper breeding strategy for developing bean varieties with durable resistance, has not been established. A study was conducted on the mode of inheritance for resistance to Psp using populations generated from crosses between three susceptible female parents (Chimbamba, Nasaka, and RC 15) and CAL 143 as a resistant male parent. The progenies were advanced to F2 and F3 generations in greenhouses at Bunda College of Agriculture and Bvumbwe Research Station. The F2 and F3 populations were evaluated for resistance to Psp in the field at Dedza and Ntchenachena. The results from both F2 and F3 populations indicated that there was a single dominant gene mode of inheritance for resistance to Psp in CAL 143. This monogenic resistance can be utilised easily and quickly in breeding for resistance to Psp. CAL 143 is, therefore, an ideal source of resistance and a valuable parent in breeding for resistance to Psp in Malawi. Key Words: Halo blight, common bean, Phaseolus vulgaris, Pseudomonas syringae, resistance genes RÉSUMÉ Le flétrissement du Halo causé par le Pseudomonas syringe pv. phaseolicola (Burkh) (Psp)) est une maladie importante du haricot commun (Phaseolus vulgaris L.) au monde. Plusieurs races de Psp existent ainsi que quelques sources de résistance dans le haricot commun ont été identifiées. CAL 143 est une lignée de CIAT améliorée du haricot commun, qui était disseminée comme une variété au Malawi et d’autres de l’Afrique australe. Elle possède une bonne résistance au Psp; cependant, le mode d’acquisition pour résistance au Psp dans CAL 143, essentiel pour dresser une stratégie appropriée au développement des variétés de haricot à résistance durable, n’a pas été établi. Une étude était conduite sur le mode d’acquisition pour résistance au Psp par l’utilisation des populations issues des croisements entre trois parents femelles susceptibles (Chimbamba, Nasaka, et RC 15) et le parent mâle résistant CAL 143. Les progénies étaient portées aux générations F2 et F3 en serre au Bunda College of Agriculture et Bvumbwe Research Station. Les populations F2 et F3 étaient évaluées pour résistance au Psp sur terrain au Dedza et Ntchenachena. Les résultats des populations F2 et F3 ont indiqué que CAL143 possédait un mode de gène dominant de résistance au Psp. Cette résistance monogénique peut être facilement et rapidement utilisée en amelioration pour résistance au Psp. CAL 143 est, par conséquent, une source idéale de résistance et un parent de valeur en amelioration pour résistance au Psp au Malawi. Mots Clés: Flétrissement du Halo, haricot commun, Phaseolus vulgaris, Pseudomonas syringae, gène de résistance INTRODUCTION Halo blight caused by Pseudomonas syringae pv phaseolicola (Psp) is a serious seed-borne disease of common bean worldwide (CIAT, 1981; Schwarts and Pastor-Corrales, 1989). It is a disease of economic importance in areas of moderate to cool temperatures in the highlands of Africa (Beebe and Pastor-Corrales, 1991). The pathogen is widely distributed in Malawi (Msuku et al., 2000) and causes serious damage in Africa (Worthmann et al., 1998). The typical symptoms are small brown spots that are surrounded by a light-green or yellow halo. The halo is caused by a toxin produced by the Psp bacterium when temperatures are less than 21oC for at least part of the day. The disease also causes water soaked greasy looking spots on pods and may appear as small, brown, scabby spots. Nine races of the pathogen have been reported based on their reactions on differential cultivars and lines (Taylor et al., 1996a). The cultivar Red Mexican U.I.3, GN Nebraska # Sel. 27 and U.I.35 have hypersensitive resistance to race 1 isolates, controlled by a single gene (Taylor et al., 1996b; Beebe and Pastor-Corrales, 1991). A recessive gene controlling tolerance to races 1 and 2 was reported in genotype P.I. 150414 from USA, while a two complementally gene model was found in a USA variety, Montcalm. A series of three recessive alleles complementally to either of two other genes were identified in a Malawian bean line 1212D (Msuku, 1984; Kelly et al., 1985). An effective breeding strategy for resistance to any disease requires a thorough understanding of the nature of inheritance of the genes controlling resistance to the pathogen causing the disease. CAL 143 has shown good resistance to Psp in Malawi, but the mode of inheritance has not been determined; hence the need for this study. The identification of additional sources of resistance to Psp and knowledge of its mode of inheritance will facilitate the incorporation of durable resistance in new improved bean varieties. The objective of this study was to determine the mode of inheritance of halo blight resistance in the common bean. MATERIALS AND METHODS Crosses were made between three recipient (susceptible) parents to Psp: Chimbamba, Nasaka, and RC 15 and one donor (resistant) parent to Psp: CAL 143. Chimbamba is a local land race, climbing bean cultivar of Type IV, which is adapted in Malawi. It is normally grown with stakes or in association with maize for support. Nasaka is a local land race, bush bean cultivar of Type I, which is adapted in Malawi, and RC 15 is also a bush bean line of Type I, which originated from the bean breeding programme at Bunda College of Agriculture in Malawi. CAL 143 is a CIAT bred line that has been released as a variety called Napilira in Malawi, where it is resistant to Psp, in addition to good adaptation as well as high yields (mean yield over 2300 kg ha-1). The variety is also grown in other countries in southern Africa region (Angola, southern Democratic Republic of Congo, Mozambique, Swaziland and Zambia). The F1s were generated and advanced to F2 and F3 in the greenhouses at Bunda College of Agriculture and Bvumbwe Agricultural Research Station. The parents, F2 and F3 populations were evaluated in the fields at Dedza and Ntchenachena sites, which represent the normal bean growing conditions during the rainy season, where halo blight is a common disease. The site in Dedza was located at 1500 meters above sea level (masl), on a geographical position: 14o 20’ S; 34o 18’ E, with annual average temperature of 20.9 °C and soils which were classified as clay loam. The site for Ntchenachena was located at 1600 masl, on a geographical position: 10o 40’ S; 34o 5’ E, with annual mean temperature of 22.4 °C and soils which were classified as clay loam. Both sites have favourable environments for Psp because of the cool temperatures and adequate wet conditions during the bean crop growing period (Msuku, 1984). The field trials were planted during the third and last week of December, respectively at Dedza and Ntchenachena. The segregating F2 and F3 progenies resulting from the crosses between the recipient parents (Chimbamba, Nasaka, and RC 15) and a common donor parent (CAL 143) were planted alongside the parental lines. Each plot had a single row of 6 meters long, and the rows were spaced at 0.75 m apart. Seeds were planted at a spacing of 10 cm for bush beans (Nasaka and RC 15) and their resulting progenies, while the climbing bean (Chimbamba) and the resulting progenies were planted at and 15 cm apart, because climbing bean plants are more vigorous and require more space. Nitrogen fertiliser was applied (40 kg N ha-1) at the first trifoliolate leaf stage and weeding was done on regular basis. Insect pests were controlled by occasional spray with Dimethoate pesticide. Rainfall data were collected during the crop growing period at both sites using rain gauges mounted at about 50 m away from the experimental fields. The individual plants were assessed based on their reaction to Psp using a CIAT scale of 1 to 9 (CIAT, 1987), where 1= immune and 9 = very susceptible. The scores were grouped into 2 categories; 1-4 for resistant plants and 5-9 for susceptible plants. The disease evaluation was based on natural occurrence of the pathogen in the trial plots. The plants were scored twice during the entire growing period: at flowering (R6) and pod filling (R8) stages. The observed disease score ratios were tested against the expected ratios through a Chi-square test using the statistical package for the social sciences (SPSS) Version 9.0. The observed phenotypic classes were tested for goodness of fit to postulated ratios based on the possible number of genes involved, and this was used to determine the mode of inheritance for resistance to Psp. The frequency distributions of plants based on disease scores were plotted to determine the overall reaction of the parental lines and segregating populations to Psp in the field. RESULTS Among the parental lines, CAL 143 was resistant to Psp, while Chimbamba showed differential reaction, and both Nasaka and RC 15 were susceptible (Fig.1, Fig. 3 and Fig. 5). In the segregating populations, a total of 17 F2 and 20 F3 plants from Chimbamba and CAL 143 cross were evaluated for resistance to Psp. The F2 population showed a bimodal distribution, while the F3 population showed a trimodal distribution pattern (Fig. 2). In the Nasaka and CAL 143 cross, a total of 21 F2 and 20 F3 plants were evaluated for resistance to Psp and both F2 and F3 populations showed a bimodal distribution pattern (Fig. 4). In the RC 15 and CAL 143 cross, a total of 25 F2 and 22 F3 plants were evaluated for resistance to Psp and both F2 and F3 populations showed a bimodal distribution pattern (Fig. 6). Overall, the results from the segregating populations indicated that all F2 populations from the three recipient parents showed a bimodal distribution pattern in their reaction to Psp . Among the F3 populations only plants from Chimbamba as a female parent segregated differently from the rest by showing a trimodal distribution pattern while those from Nasaka and RC 15 showed a bimodal distribution pattern in reaction to Psp . Inheritance of resistance to Psp.The data collected from the evaluation of the F2 and F3 populations, were arranged in two categories: resistant (disease scores 1-4) and susceptible (disease scores 5-9) based on their reaction to Psp . These data were analysed using a Chi-square test. The Chi-square additivity test on both F2 and F3 progeny plants suggested that the data were homogeneous, X2 additivity = 0.465, P= 0.793 the resistant: susceptible ratio of 3:1 at F2 and 5:3 at F2 and X2 additivity = 0.181, P= 0.913 at F3 (Table at F3; indicating that CAL 143 had one dominant 1). The segregation ratios were, therefore, tested gene conferring resistance to Psp (X2 pooled= based on pooled Chi-square and the results 1.01, P= 0.799 for F2 and X2 pooled= 1.722, P= 0.632 indicated that the data were in compliance with for F3). DISCUSSION The findings from this study suggest that it is practically possible to phenotype individual plants for resistance to Psp under natural conditions as long as the pathogen is present and the environmental conditions are favourable. Asensio et al. (2010) conducted field evaluations under inoculated conditions and identified two accessions (BGE 002189 and BGE 029592) out of 199 from a Spanish core collection that had immune reaction to two races of Psp (race 6 and 7). Msuku (1984) suggested that field tests would be useful in detecting field resistance more accurately than greenhouse or laboratory tests. The results from this study also suggest that there is one dominant gene conferring resistance to Psp in CAL 143 (Table 1). This mode of inheritance of resistance to Psp, which is controlled by a single dominant gene, was also reported by Miklas et al. (2009). They conducted allelism tests involving UI-3 (formerly ‘Red Mexican UI-3’) and host differential cultivars: Canadian Wonder, ZAA 54 (A52), Tendergreen, ZAA 55 (A53), ZAA 12 (A43) and Guatemala 196B, where a single dominant gene, Pse-1, was reported to condition resistance to races 1, 7, and 9 of Psp. Thus the single dominat gene was found to be a valuable source of broad spectrum resistance to Psp. This gene has been mapped to linkage group B 10 and found to be near quantitative trait loci (QTL) for resistance to fusarium wilt, halo blight, and angular leaf spot (Fall et al., 2001; Blair et al., 2003; Lopez et al., 2003; Miklas et al., 2006). In a study conducted by Msuku (1984), two dominant complementary genes were found to determine resistance to race 2 of Psp in Montcalm. In the same study, a Malawian land race, 1212D had an allelomorphic series of three alleles, which were assumed to control resistance to the same race in complement with either of two genes. Source of broad resistance. Although in this study, the actual number of prevalent halo blight races in the field were not determined, Msuku (1984) identified four races (1, 2, 3 and 4) after screening 32 isolates, which were obtained from various bean growing areas of Malawi, including Dedza and Nchenachena. These races were identified based their reaction to five differential cultivars (Red Mexican UI-3, Red Mexican UI-34, Great Northern 123, Jubila and Namajengo (a landrace in Malawi). Nasaka and Montcalm were used as susceptible and resistant controls. Many of the isolates belonged to race 2 (16 isolates) and race 1 (13 isolates), indicating that these races might be common in most bean production environments where Psp is predominant. Since Msuku’s (1984) results suggested more than one race of Psp in the bean production environments, it would appear that CAL 143 may potentially offer a good source of resistance to a broader range of Psp races, which are prevalent in Dedza and Ntchenachena, the selected two major bean growing areas in this study where Psp is predominat. That a single gene can offer broad resistance to Psp was demonstrated by Miklas et al. (2009), and this was found in the differential cultivar UI-3. Teverson (1991) and Taylor et al. (1996b), have reported five putative R genes (R1, R2, R3, R4, and R5) that have been identified to confer resistance to Psp, and the gene action, dominant or recessive are symbolised Pse or pse respectively (Basset, 2004). Fourie et al. (2004) reported that genes conferring resistance to Psp occur in a tight cluster. CAL 143 has shown to potentially have Pse gene which confers broad resistance to Psp, at least against the races that might occur across the two bean production environments (Dedza and Nchenachena) in Malawi. Asensio et al. (2010) also identified two accessions from a Spanish core collection that had resistance to Psp races 6 and 7, which have not been reported in Malawi. Implications for breeding. The pooled Chi-square value for F2 populations strongly supported a single gene model of inheritance in CAL 143 to Psp. The F3 data were equally supportive and confirmed the same model. CAL 143, with broad resistance to Psp, is already released as a variety in Malawi, because of high yield and its attractive grain type for the market (medium to large seed size and red-mottled grain colour). The variety can be widely promoted for production in the areas where Psp is a constraint to bean production. In addition, CAL 143 can be utilised in the breeding programmes when making future crosses for developing new varieties with high yield and resistance to Psp. The use of a broad genetic base has been advocated by Singh (2001) as a good strategy for varietal improvement. The fact that its resistance is monogenic and dominant makes the breeding process easier and quicker to make progress and achieve the intended breeding objectives. CONCLUSION The inheritance of resistance to Psp in CAL 143 is determined by a single dominant gene (Pse). This genotype is an ideal source of resistance to Psp in Malawi and a good parent in breeding for resistance to Psp in common beans. It would be easier to breed desirable bean varieties with resistance to Psp using CAL 143 for Malawi and other countries in southern Africa, where the preference is for large seeded (Andean) bean types, because the resistance gene is already in the large seeded background and it is determined by a single dominant gene. ACKNOWLEDGEMENT We are grateful to the Bean/Cowpea CRSP for funding this research and to CIAT/Pan-Africa Bean Research Alliance (PABRA) for providing germplasm for the source of resistance to Psp. We also thank staff from the Bean-Cowpea project at Bunda College of Agriculture, Lunyangwa and Bvumbwe Agricultural Research Stations and Nchenachena Agricultural Research sub-Station for providing technical support. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11030f1.jpg] [cs11030f4.jpg] [cs11030f3.jpg] [cs11030f5.jpg] [cs11030f6.jpg] [cs11030t1.jpg] [cs11030f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}