 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
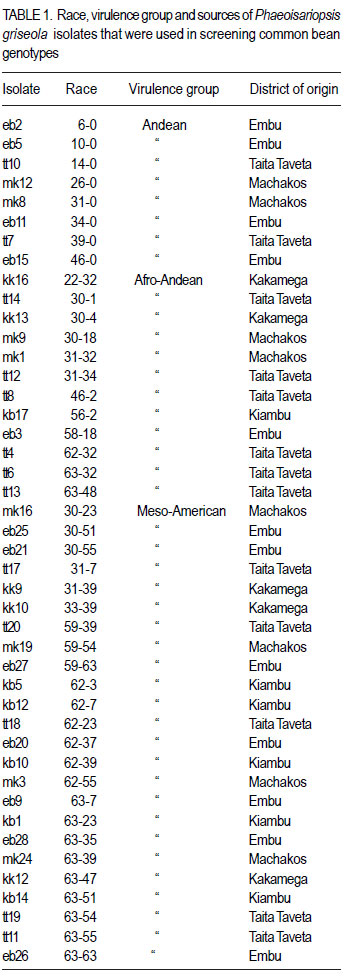
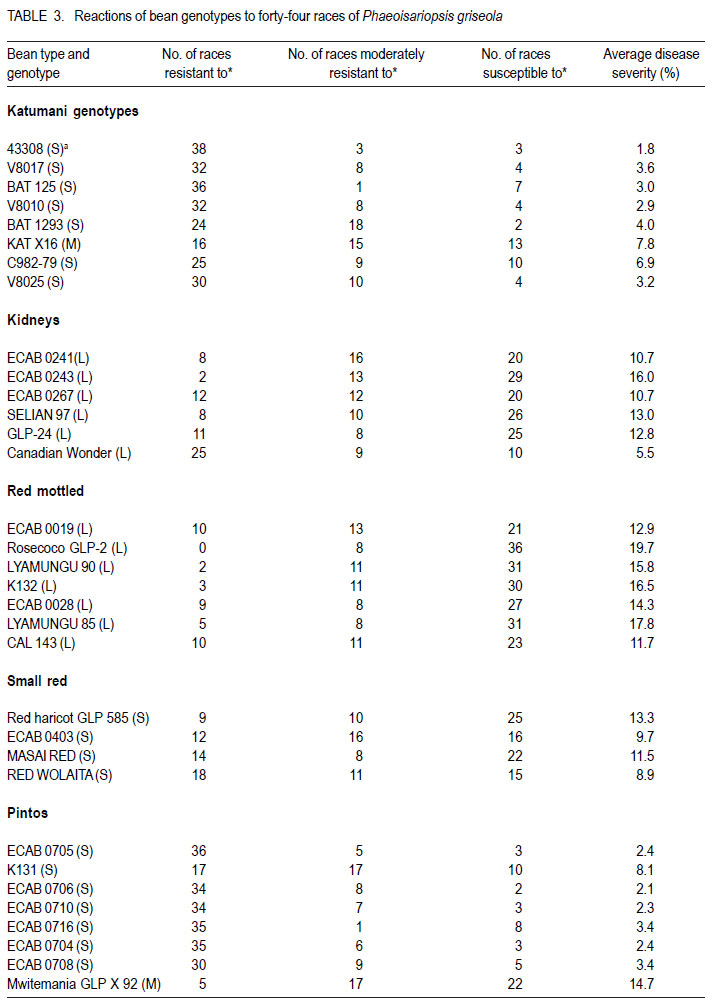
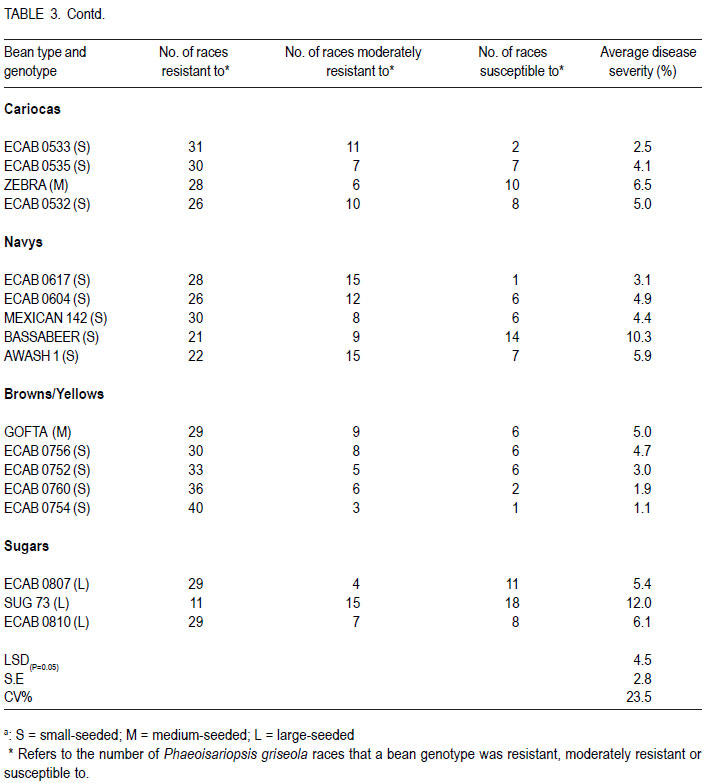
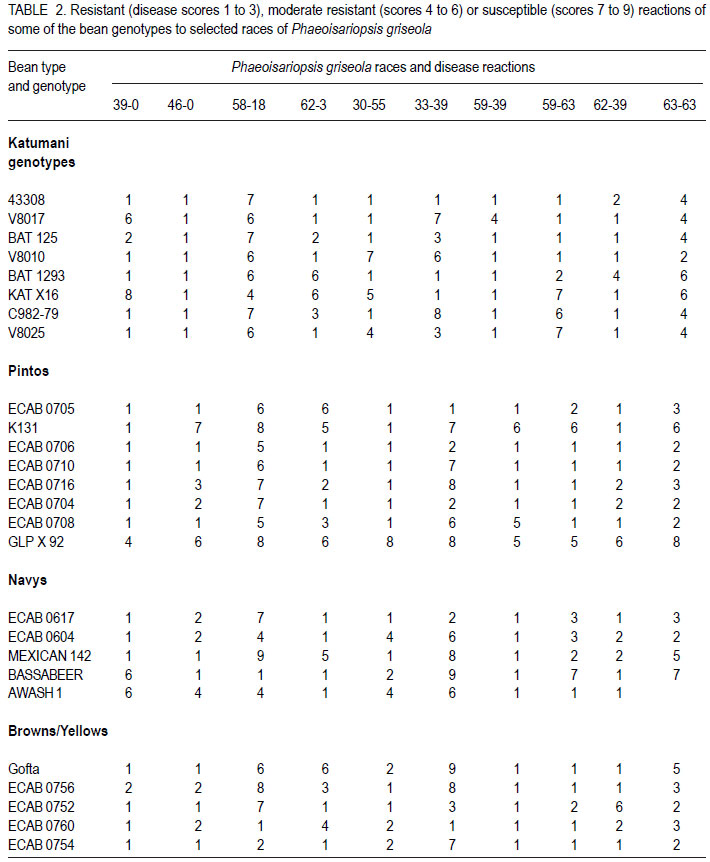
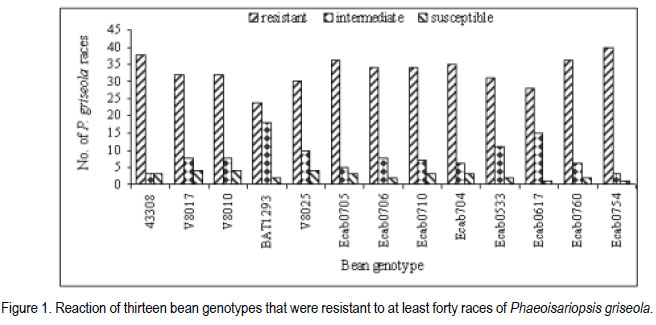
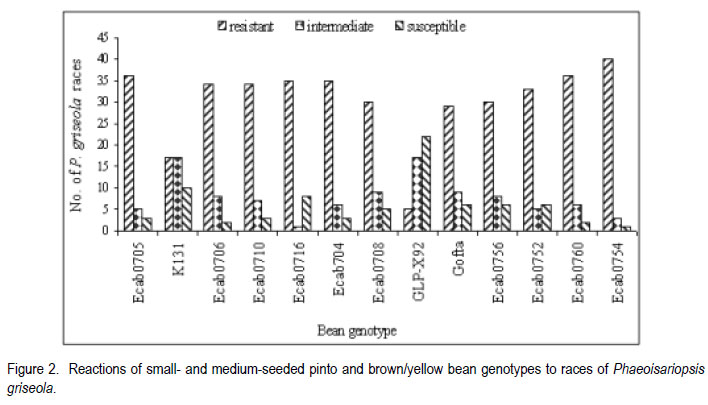
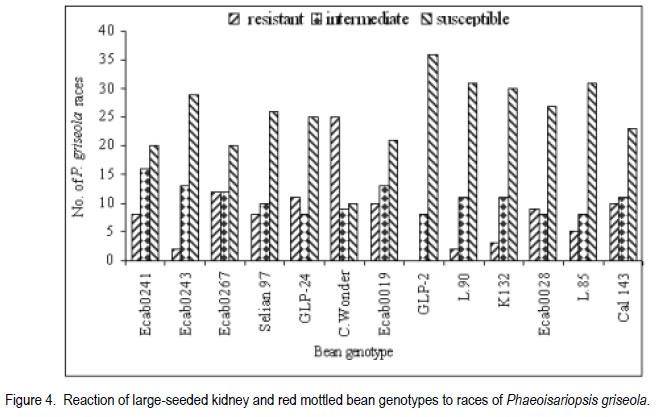
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 343-355 Reaction of selected common bean genotypes to physiological races of Phaeoisariopsis griseola occurring in Kenya I.N. Wagara, A.W. Mwang’ombe1, J.W. Kimenju1, R.A. Buruchara2 and P.M. Kimani Department of Biological Sciences, Egerton University, P. O. Box 536, Egerton, Kenya ABSTRACT The wide pathogenic variability occurring in Phaeoisariopsis griseola, the causal agent of angular leaf spot of common bean (Phaseolus vulgaris L.), dictates that new sources of resistance be continuously identified. This study was undertaken to determine the reaction of selected bean genotypes to different races of P. griseola in order to identify potential sources of resistance to angular leaf spot. Selected bean genotypes from Eastern and Central Africa Bean Research Network (ECABREN) and National Dryland Farming Research Centre (NDFRC), Katumani in Kenya were separately inoculated with forty-four races of P. griseola and evaluated for disease development under greenhouse conditions. The genotypes included small- and large-seeded types. None of the genotypes was resistant to all the races, indicating a high complexity of the pathogen population. Thirteen genotypes were resistant (disease score 1 to 3) or moderately resistant (score 4 to 6) to at least 40 of the races. Small-seeded bean genotypes ECAB 0754 and ECAB 0617 were resistant or moderately resistant to all races except Mesoamerican race 33-39 and Afro-Andean race 58-18, respectively. Genotype ECAB 0754 exhibited the highest level of resistance, with an average disease severity of 1.1%. All the resistant or moderately resistant genotypes were of the small-seeded bean types which are commercially less popular. The commonly grown large-seeded genotypes were generally susceptible. Among the bean genotypes evaluated, the small-seeded pintos and browns/yellows possessed high levels of resistance. The results of this study indicate that different bean genotypes have varying levels of resistance to angular leaf spot that can be pyramided into appropriate background to provide durable resistance. Key Words: Angular leaf spot, Pathogenic variability, Phaseolus vulgaris RÉSUMÉ Une large variabilité pathogénique du Phaeoisariopsis griseola, l’agent causal de la tâche angulaire de feuilles du haricot commun (Phaseolus vulgaris L.), suggère qu’il y’ait une identification continue de nouvelles sources de résistance. Cette étude était entreprise pour déterminer la réaction des génotypes de haricot sélectionnés aux différentes races de P. griseola afin d’identifier de sources potentielles de résistance à la maladie de tâche angulaire de la feuille. Les génotypes de haricot fournis par l’Eastern and Central Africa Bean Research Network (ECABREN) et le National Dryland Farming Research Centre (NDFRC), Katumani au Kenya étaient séparément inoculés avec quarante-quatre races de P. griseola pour l’évaluation en serre du développement de maladies. Les génotypes comprenaient de petits et gros grains de semences. Aucune des génotypes n’était résistant à toutes les races de maladies, indicant ainsi une complexité élevée de la population des pathogènes. Treize génotypes étaient résistants (échelle de cotation: 1 à 3) ou modérément résistants (échelle de cotation: 4 à 6) sur au moins quarante de toutes les races. Les génotypes de haricot à petits grains ECAB 0754 et ECAB 0617 étaient résistants ou modérément résistants à toutes les races à l’exception de Mesoamerican race 33-39 et Afro-Andean race 58-18, respectivement. Le Génotype ECAB 0754 avait manifesté un niveau élevé de résistance, avec un taux moyen de séverité de 1.1%. Tous les génotypes résistants ou modérément résistants étaient de ceux à petits grains commercialement moins populaires. Les génotypes à gros grains communément cultivés étaient généralement susceptibles. Parmi les génotypes évalués, ceux à petits grains “pintos”et bruns/jaunes étaient dotés des niveaux élevés de résistance. Les résultats de cette étude montrent que différents génotypes de haricot possèdent de niveaux différents de résistance à la maladie de tâche angulaire de la feuille pouvant être combinés par pyramidage pour développer une variété de haricot à résistance durable. Mots Clés: Tâche angulaire de la feuille, Variabilité pathogénique, Phaseolus vulgaris INTRODUCTION Common bean (Phaseolus vulgaris L.) is widely cultivated in tropical and subtropical countries, where it is an important source of proteins, carbohydrates, fiber, vitamins and minerals (Wortmann et al., 1998; Voysest, 2000). In Kenya, common bean is the most important legume in the pulses category and is second only to maize as a food crop (Gethi et al., 1997). Despite its importance, bean yields in developing countries are among the lowest in the world, with average of 0.5 tonnes ha-1 (FAO, 2007) compared to 1–2 tonnes ha-1 commonly reported in experimental fields. The low yields are attributed to a number of constraints, most important of which are diseases, insect pests, low soil fertility and periodic water stress (Allen et al., 1989; Otysula et al., 1998). Angular leaf spot (ALS) of common bean, caused by Phaeoisariopsis griseola (Sacc.) Ferraris, is one of the most damaging and widely distributed diseases of common bean, causing yield losses as high as 80% (Stenglein et al., 2003). In Africa, particularly in Kenya, Malawi, Ethiopia, Uganda, Tanzania, where beans constitute the most important source of dietary protein, ALS is a significant constraint causing annual losses estimated at 374,800 tonnes in the region (Wortmann et al., 1998). Use of resistant varieties is pivotal to any effective, economical and environmentally friendly strategy of managing ALS, especially for small-scale farmers in sub Saharan Africa. The advantage of host plant resistance is that once the technology has been developed, it is packaged in seed which is easier to disseminate and deploy, and does not require any additional or specialised handling on the part of the farmers (Mahuku et al., 2009). The greatest setback to development and deployment of resistant bean varieties is the high pathogenic variability occurring in P. griseola that renders varieties that are resistant in one location or year susceptible in another (Mahuku, et al., 2002a). Characterisation based on differential cultivars, random amplified polymorphic DNA (RAPD) markers, microsatellites, isozymes and amplified fragment length polymorphism (AFLP) have revealed high levels of pathogenic and genetic variation in P. griseola (Guzman et al., 1999; Mahuku et al., 2002a; 2002b; Wagara et al., 2004; 2005). In Kenya, forty-four physiological races of P. griseola have so far been identified (Wagara et al., 2005). Despite their great diversity within and between countries, isolates of P. griseola have been classified into Andean, Afro-Andean and Mesoamerican (Mahuku et al., 2002a; 2002b; Wagara et al., 2005). Andean isolates of P. griseola do not infect Mesoamerican beans, while Mesoamerican isolates infect beans from both gene pools but are more virulent on Mesoamerican beans. Afro-Andean isolates, on the other hand, infect one or two Mesoamerican differential genotypes in addition to the Andean. Resistance genes of Mesoamerican origin are more effective in managing Andean ALS pathotypes and vice versa (Mahuku et al., 2009). Inheritance studies have shown that ALS resistance in common bean is mediated by both major and minor genes that are either dominant or recessive with complementary or epistatic effects and acting alone or in combination (Carvalho et al., 1998; Mahuku et al., 2004; Caixeta et al., 2005). Sartorato (1989) reported that some bean varieties (Cornell 49242, RG1342, CH60, Mexico 279 and Mexico 54) possess high levels of race-specific or vertical resistance, while others like Compuesto Chimaltenango 2 have race non-specific or presumed horizontal resistance to angular leaf spot. There are also indications that rate-reducing resistance against P. griseola occurs in some bean varieties whereby varieties show differences in the time when symptoms appear and the extent of disease severity (Buruchara, 1983; Wagara, 1996). Bean cultivars that are resistant to some P. griseola races are available (Liebenberg and Pretorius, 1997; Pastor-Corrales et al., 1998; Ferreira et al., 2000; Stenglein et al., 2003), but there are no genotypes that are resistant to all the pathogen races. Thus, better sources of multiple resistance to ALS should be identified. The high pathogenic variability of P. griseola population and the wide distribution of the pathogen races imply that using a single source of resistance would be insufficient in management of ALS. Pyramiding several resistance genes in one variety is one way of breeding broad and durable resistance into dry bean (Mahuku et al., 2002b). Combining Andean and Mesoamerican resistance genes into the same background is more likely to result in durable resistance to many ALS pathotypes (Mahuku et al., 2009). An important step towards achieving this goal involves screening of the available germplasm against a broad range of P. griseola population. This study was, therefore, undertaken to determine the response of selected bean genotypes to races of P. griseola occurring in Kenya in an effort to identify potential sources of resistance to ALS. MATERIALS AND METHODS Source of Phaeoisariopsis griseola isolates. Forty four isolates of P. griseola, which were representative of 44 physiological races of the pathogen identified in Kenya, were used in this study (Table 1). Eight of the races were Andean, 12 Afro-Andean and 24 were Mesoamerican. The isolates had been obtained from naturally ALS infected bean leaves collected from farmers’ fields in different bean growing areas in Kenya, and characterised into races based on their reactions on the twelve international differential bean cultivars (Wagara et al., 2005). Selection and inoculation of bean genotypes. A total of two hundred bean genotypes were used in this study. One hundred and fifty two genotypes were from Eastern and Central Africa Bean Research Network (ECABREN), 40 from National Dryland Farming Research Centre (NDFRC) at Katumani, and eight advanced lines from the Bean Breeding Programme of the University of Nairobi. The bean genotypes included both small- and large-seeded types and were selected based on yield performance and/ or resistance to diseases such as bean anthracnose and rust. A preliminary evaluation of the two hundred bean genotypes for reaction to the most virulent race of P. griseola (Mesoamerican race 63-63) was undertaken and the results were used to select genotypes for further evaluation with the other forty-three races of the pathogen. Reactions of the bean genotypes to races of P. griseola were assessed under greenhouse conditions. Bean seeds were surface-sterilised in 2.5% sodium hypochlorite for three minutes and rinsed in five changes of sterile distilled water. The seeds were sown in sterile soil composed of soil, sand, manure and ballast in the ratio of 2:1:1:1. Five seeds of each genotype were sown per two kilogramme pot capacity and thinned to three plants two weeks after sowing. Three pots (a total of nine plants) were used for each genotype. Monosporic cultures of P. griseola were grown on bean leaf dextrose agar (BLDA) for 14 days, in a non-illuminated incubator at 24 oC. Inoculum was prepared by adding sterile distilled water onto culture plates and scrapping the culture surface with a sterile glass rod. Three-weeks-old seedlings were spray-inoculated with the pathogen at a concentration of 2x104 conidia ml-1 until runoff, using a hand-sprayer from East Africa Seed Company. Plants sprayed with sterile water were included as the control. The plants were placed on a greenhouse bench and maintained in high humidity through frequent mist sprays to facilitate disease development. A completely randomised design with nine replicates was used. Disease assessment. Inoculated plants were examined daily for ALS symptom development, up to 21 days. On appearance of ALS symptoms, plants were scored for disease severity four times after every three days using a scoring scale of 1 to 9 (Van Schoonhoven and Pastor-Corrales, 1987). Reaction type categories were determined from the average of disease scores for each isolate-host genotype combination. Plants showing disease scores 1 to 3 were considered resistant, 4 to 6 as moderately resistant and host responses corresponding to scores 7 to 9 as susceptible. Forty six resistant or moderately resistant bean genotypes were selected for further evaluation with the other forty three races of P. griseola occurring in Kenya. Four of the commonly grown bean varieties (Rosecoco GLP-2, Mwitemania GLPX.92, Red Haricot GLP-585 and Canadian Wonder GLP-24) were included as checks. Each race of the pathogen was handled in a separate inoculation chamber to avoid cross-infection. The experiment (planting, inoculation and disease evaluation) was repeated once to confirm results. Data on percent disease severity of each isolate-host genotype interaction were analysed by the analysis of Variance (ANOVA) using GenStat Release 6.1 (GenStat Procedure Library Release PL14). Separation of means was done using least significant difference (LSD) test at P < 0.05. RESULTS Out of the 200 bean genotypes inoculated with P. griseola race 63-63, fourteen (7%) were resistant (disease scores 1 to 3), sixty-seven (33.5%) were moderately resistant (scores 4 to 6) and 119 (59.5%) were susceptible (data not shown). The fifty bean genotypes selected for further evaluation with all the forty four races of the pathogens exhibited varying reactions and the differences in disease severity were significant (P<0.05) (Table 3a, 3b). None of the genotypes was resistant to all races of the pathogen. Some genotypes, however, were either resistant (scores 1 to 3) or moderately resistant (scores 4 to 6) to most of the races and only susceptible (scores 7 to 9) to a few races (Table 2). A few genotypes exhibited high levels of resistance or immunity to most of the races. For example, bean line ECAB 0754 was resistant to 40 races of the pathogen (Table 3a, 3b). Eight of the genotypes (43308, ECAB 0533, ECAB 0704, ECAB 0705, ECAB 0706, ECAB 0710, ECAB 0754 and ECAB 0760) had an average severity ranging from 1.1 to 2.5% (scores1 to 3) and were considered to be resistant (Table 3a, 3b). Twenty five genotypes had an average severity score of more than 2.5, but < 7% and were considered to be moderately resistant; whereas 17 genotypes were susceptible. Bean line ECAB 0754 had the lowest average disease score (1.1%), whereas the popular bean variety, Rosecoco GLP2 had the highest score of 19.7% (Table 3a, 3b). Thirteen genotypes were resistant or moderately resistant to at least 40 of the races (Fig. 1). All the resistant or moderately resistant genotypes were of the small-seeded bean types, whereas the popular large-seeded beans were generally susceptible (Table 3a, 3b). Among the various bean categories evaluated, pintos (small-seeded) had the highest number of resistant genotypes (Table 3a, 3b). Out of the eight pinto bean genotypes tested, six were resistant or moderately resistant to at least 35 (78%) of the races (Fig. 2). Bean lines ECAB 0705, ECAB 0706, ECAB 0710 and ECAB 0704 (all small-seeded) were resistant or moderately resistant to more than 40 (91%) of the races. Mwitemania GLP-X.92, a popular medium-seeded pinto bean variety, was susceptible to most of the races and only exhibited resistance to five races (Table 3a, 3b). The small-seeded brown/yellow bean types also exhibited high levels of resistance and all the five genotypes tested from this category were resistant to at least 38 races (Fig. 2). Of these five genotypes, ECAB 0754 was resistant or moderately resistant to all races except Mesoamerican race 33-39, whereas the least resistant genotypes (Gofta, ECAB 0756 and ECAB 0752) were susceptible to only six races each (Table 3a, 3b). Among the sugars category (largeseeded types), ECAB 0807 and ECAB 0810 were resistant to 29 races each and susceptible to 11 and 8 races, respectively (Table 3a, 3b). All the carioca and navy genotypes tested were resistant to at least 30 races. Navy bean genotype ECAB 0617 (small-seeded) was resistant or moderately resistant to all races except race 58-18, whereas ECAB 0533 (small-seeded carioca) was susceptible to only two races. In these two bean categories, Bassabeer was the most susceptible, succumbing to 14 races of the pathogen. The small-seeded bean genotypes from NDFRC Katumani that were tested in this study showed high levels of resistance to different races of P. griseola (Fig. 3). All the eight genotypes were resistant or moderately resistant to at least 30 (68%) of the races. Five of the genotypes (43308, V8010, V8017, V8025 and BAT1293) were resistant to at least 40 races, with an average ALS severity score of less than 3.6% (Table 3a, 3b). Bean genotypes BAT 1293 and 43308 were susceptible to only two and three races, respectively whereas V8017, V8010 and V8025 succumbed to four races each. Only three of the genotypes (medium-seeded KAT X16, small-seeded BAT 125 and small-seeded C982-79) were susceptible to more than five races. The commonly grown large-seeded bean genotypes (kidneys and red mottled) were generally susceptible (Fig. 4). The most popular variety, Rosecoco GLP-2 (large seeded) was susceptible to most of the races with an average severity score of 19.7% and was only moderately resistant to eight races. Canadian Wonder GLP-24 was the best performing largeseeded variety, being resistant or moderately resistant to 34 races. The small-red bean types also showed high levels of susceptibility to most races of P. griseola (Table 3a, 3b). Red Haricot GLP 585, a popular small-seeded red bean, was resistant or moderately resistant to only 19 races of the pathogen. DISCUSSION This study revealed a wide variation in the reactions of bean genotypes to the forty four races of P. griseola occurring in Kenya. None of the genotypes evaluated was resistant to all races of the pathogen, indicating a high complexity of the pathogen population. However, thirteen genotypes were resistant or moderately resistant to at least 40 of the races. Bean genotypes, ECAB 0754 and ECAB 0617, were resistant or moderately resistant to all races except Mesoamerican race 33-39 and Afro-Andean race 58-18, respectively. These genotypes are, therefore, good sources of resistance to ALS and can be complimented with genes from other resistant genotypes to buildup durable resistance. For example, resistance to race 33-39 and race 58-18, the only races that induced susceptible reactions in genotypes ECAB 0754 and ECAB 0617, respectively, can be obtained from ECAB 0760 which showed high levels of resistance to these two races. All the bean genotypes showing high levels of resistance to angular leaf spot were of the small-seeded types with very little resistance among the popular large-seeded genotypes, indicating the importance of genepool type in the search for resistance to ALS. The popular large-seeded genotypes were generally susceptible, with Roseccoco GLP-2 (the commonly grown variety in Kenya) being the most susceptible. These results are consistent with the report by Pastor-Corrales et al. (1998) who observed that of the 19 bean accessions that were intermediate or resistant to ALS, 15 were small or medium-sized while only four were large-seeded. These results are also in support of earlier observations from CIAT (1996) that the resistant bean accessions, often without any visible ALS symptoms, are the small-seeded genotypes of Middle American origin. Liebenberg and Pretorius (1997) also reported that most of the known sources of ALS resistance are evidently small-seeded bean varieties and that very little resistance exists among the popular large-seeded varieties in Southern Africa. One way of circumventing the lack of resistance in well-adapted bean varieties of market types preferred by farmers and consumers would be to pyramid complementary resistance genes into these varieties. Pyramiding complementary resistance genes into appropriate backgrounds is a strategy with potential for maintaining long-term durable disease resistance (Young and Kelly, 1996; Mahuku et al., 2009). This has been demonstrated in bean anthracnose differential genotype G2333, which contains three resistance genes and only one pathotype of Colletotrichum lindemuthianum has been found to infect it (Pastor-Corrales et al., 1994; Young et al., 1998). The rationale behind gene stacking is that combining as many resistance genes in the same background as is practical, makes it statistically very difficult for a pathotype to evolve, that would overcome all the resistance genes simultaneously (Mahuku et al. 2009). Among the bean types evaluated in the present study, the small- to medium-seeded were found to possess high levels of resistance to ALS. These bean types are less popular with farmers and consumers in Kenya but can be used as sources of resistance to improve the popular, but susceptible, varieties. Bean genotypes identified in this study to possess varying levels of resistance can be promoted for inclusion in farmers’ varietal mixtures to boost management of ALS. Pyndji and Trutmann (1992) demonstrated that ALS could be controlled in farming systems where local bean mixtures are used by supplementing them with resistant varieties. Wolfe (1985) noted that host mixtures may restrict the spread of pathogens and diseases considerably, provided the varieties differ in their susceptibility. Use of an assortment of the resistant genotypes by farmers would ensure the presence of different sources of resistance to ALS in a region. The physical presence of a resistant variety would provide farmers with some assurance of yields during a favourable growing season. It has been shown that adding high yielding ALS resistant varieties to local farmer bean mixtures can result in dry bean yields comparable to those obtained from the high yielding resistant variety cultivated alone (Trutmann and Pyndji, 1994). Thus, farmers should be encouraged to diversify their bean varietal types to also include the currently commercially less popular seed types that may possess resistance to angular leaf spot. Bean genotypes identified in this study under greenhouse conditions as potential sources of resistance should be tested at various angular leaf spot hot spots to establish their levels of resistance under field conditions. Field sites with the highest P. griseola population diversity should be selected where the varieties can be tested against the widest range of natural inoculum. The wide pathogenic variability in the ALS pathogen population in Kenya implies that using a single location to test varieties for resistance is not sufficient because different races occur in different locations (Wagara et al., 2005). Because none of the bean genotypes evaluated in this study was resistant to all races of P. griseola, screening of more bean germplasm should be intensified in an effort to identify new and better sources of resistance to ALS. Due to the high pathogenic variability in the ALS pathogen, it is desirable to use broader sources of resistance. The fact that eight and five of the bean genotypes obtained from ECABREN and NDFRC Katumani, respectively, were resistant or moderately resistant to most races of P. griseola indicates that screening of more genotypes from these and other bean improvement programmes would be useful. Evaluations by Pastor-Corrales et al. (1998) revealed that 89 bred bean lines were resistant or intermediate resistant despite the fact that only two (MAR 1 and MAR 3) were deliberately bred for ALS resistance. Screening should especially target bean genotypes of the small-seeded genepool which has been shown to possess good levels of resistance to ALS (CIAT, 1996). It would also be worth while to evaluate the various common bean landraces grown by farmers in different regions for resistance. Germplasm screening for ALS resistance should also be extended to wild common bean populations, which may harbour sources of resistance. Screening of other Phaseolus spp. such as P. costaricensis, P. coccineus, P. polyanthus, P. acutifolius and P. parvifolius that can be hybridised with common bean has been recommended (Pastor-Corrales et al., 1998). High levels of resistance to ALS have been found in P. coccineus and P. polyanthus (Mahuku et al., 2003), confirming earlier reports that the secondary gene pool is an important sources of useful traits (Bosogoro et al., 1999). Sources of resistance to other diseases and pests (e.g. resistance to Aschochyta, common bacterial blight, and bean golden mosaic virus) have been found in the secondary gene pools (Beebe and Pastor-Corrales, 1991). The effective strategy of breeding for ALS resistance would be to hybridise those bean varieties that possess broad and complementary genes for resistance. This should be followed by field screening and selection in sites where the widest range of P. griseola populations are endemic. ALS resistance lines thus developed would then be tested in the greenhouse against a more diverse group of isolates that cannot, for various reasons, be used in field inoculations. ACKNOWLEDGEMENT Financial support for this work was provided by The Rockefeller Foundation (Grant 2001 FS 030). We gratefully acknowledge the Pan-Africa Bean Research Alliance (PABRA) through its Eastern and Central Africa Bean Research Network (ECABREN) and National Dryland Farming Research Centre (NDFRC) Katumani for providing the bean genotypes used in this study. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11032f2.jpg] [cs11032f4.jpg] [cs11032t3.jpg] [cs11032f1.jpg] [cs11032t1.jpg] [cs11032t3b.jpg] [cs11032f3.jpg] [cs11032t2.jpg] [cs11032t3a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}