 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 369-376 Application of molecular markers in breeding for bean common blight resistance in South Africa D. Fourie and L. Herselman1 Agricultural Research Council-Grain Crops Institute, Private Bag X1251, Potchefstroom, 2520, South Africa
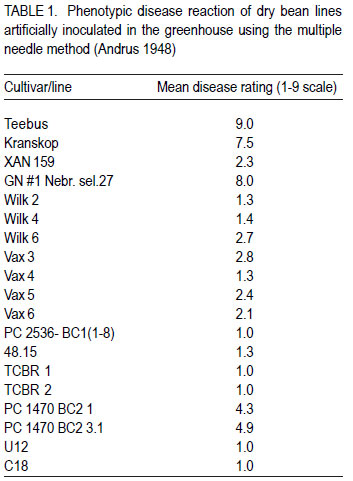
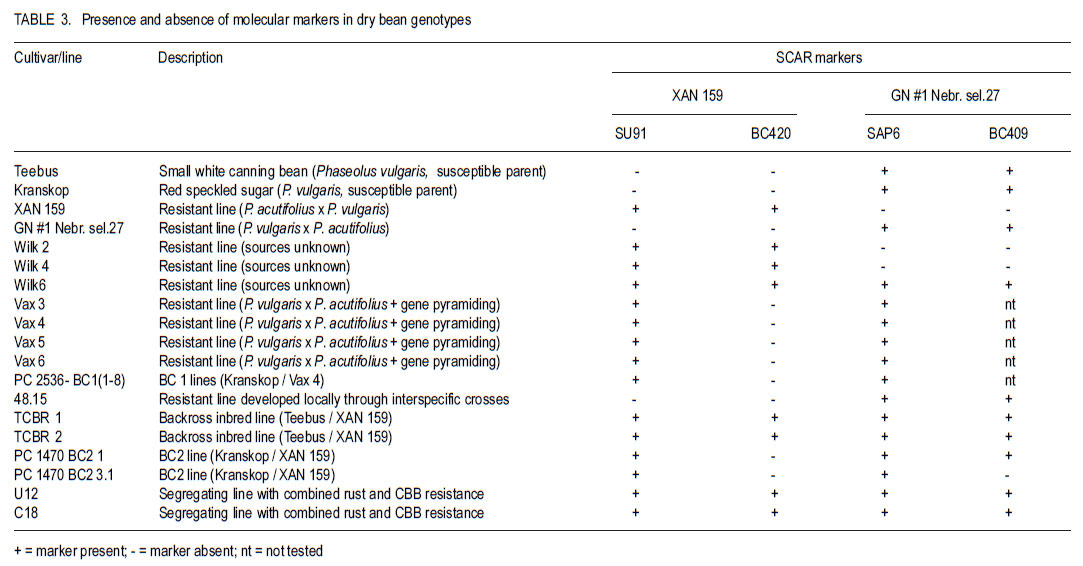
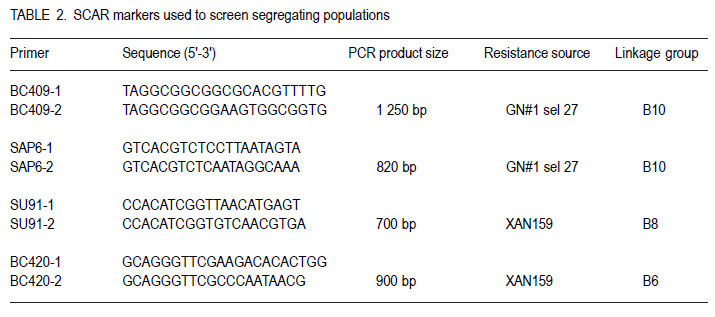
Code Number: cs11034 ABSTRACT Sequence characterised amplified region (SCAR) markers, linked to four independent quantitative trait loci (QTL) in XAN 159 and GN #1 Nebr. sel. 27, are available for indirect selection of resistance to common bacterial blight in Phaseolus vulgaris. Existing SCAR-markers, SU91, BC420, BC409 and SAP6, were evaluated for potential use in the South Africa breeding programme. Segregating populations of progenies developed through backcross breeding with cultivars Teebus and Kranskop as susceptible recurrent parents, and XAN 159 and Vax 4 as resistant donor parents were evaluated for presence of existing markers. Presence of all four markers in improved Teebus lines (XAN 159 derived) confirmed transfer of resistance in these lines. Marker BC420 was absent in XAN 159 derived Kranskop-lines. These lines were only moderately resistant when tested in the greenhouse, indicating that the QTL linked to this marker is important in order to obtain high levels of resistance. Progenies from backcrosses with Kranskop as recurrent parent using Vax 4 have exhibited high levels of resistance and presence of all markers found in Vax 4 confirmed transfer of resistance. Marker-assisted selection can successfully be implemented in breeding for common bacterial blight resistance in South African materials. Key Words: Marker-assisted selection, Phaseolus vulgaris, Xanthomonas axonopodis RÉSUMÉ Les marqueurs de régions amplifiées de séquences caractérisées (SCAR), associées à quatre traits de loci quantitatives indépendantes (QTL) dans XAN 159 et GN #1 Nebr. sel. 27, sont disponibles pour une sélection indirecte de résistance au flétrissement commun dans Phaseolus vulgaris. Les marqueurs existants SCAR, SU91, BC420, BC409 et SAP6, étaient évalués pour leur utilisation potentielle dans le programme d’amélioration en Afrique du Sud. Les populations ségrégantes des progénies développées à travers l’amélioration par croisement en retour avec les parents susceptibles recurrents et les parents donneurs résistants XAN 159 et Vax 4 étaient évaluées pour la présence des marqueurs existants. La presence de tous les quatre marqueurs dans les lignées de Teebus améliorées (XAN 159) a confirmé le transfert de résistance dans ces lignées. Le marqueur BC420 était absent dans XAN 159 dérivé des lignées Kranskop. Ces lignées étaient seulement modérément résistantes lorsqu’ elles étaient testées en serre, indiquant l’importance du QTL associé à ce marqueur dans l’obtention de niveaux élevés de résistance. Les progénies issues de croisements en retour avec Kranskop comme parent recurrant utilisant Vax 4 ont montré des niveaux élevés de résistance, et la présence de tous les marqueurs trouvés dans Vax 4 ont confirmé le transfert de résistance. La sélection par marqueur asssisté peut être appliqué avec succès dans le programme d’amélioration pour résistance au flétrissement bactérien commun dans le matériel Sud Africain. Mots Clés: Sélection par marqueur assisté, Phaseolus vulgaris, Xanthomonas axonopodis INTRODUCTIONReliable methods are important in resistance breeding for phenotypic selection of resistant plant progenies. Screening for resistance may be limited when breeders are challenged with mixed races of a pathogen or when many resistance genes are present in the host (Kelly and Miklas, 1999). Environmental conditions can influence disease development in the field and the possibility of escapes also exists. Indirect selection for resistance using molecular markers offers breeders a viable alternative to confirm presence of favourable gene combinations in new breeding lines (Kelly and Miklas, 1999). Molecular markers linked to common bacterial blight (CBB) resistance in dry beans (Phaseolus vulgaris L.) have been developed for indirect selection of resistance to Xanthomonas axonopodis pv. phaseoli (Smith) Vauterin, Hoste, Kosters and Swings, (Bai et al., 1996; Beattie et al., 1998; Park et al., 1999; Yu et al., 1999; Miklas, 2002). Resistance sources GN #1 Nebr. sel 27 and XAN 159 each contribute two independent quantitative trait loci (QTL) with major effects on CBB resistance (Miklas et al., 2000). Sequence characterised amplified region (SCAR) markers (SU91, BC420, BC409 and SAP6) linked to these four QTL are available for DNA marker-assisted breeding (Miklas et al., 2000). Advanced cranberry, pinto, Great Northern and snap bean germplasm with combined resistance to common blight has been developed in the USA using these markers in the selection process (Miklas et al., 2000; Mutlu et al., 2005 Mutlu et al., 2008). SCAR marker, SU91 is, however, most frequently used in breeding programmes (Miklas et al., 2006) Yu et al. (1999) screened 138 F5 lines derived from HR67 (resistance derived from XAN 159), using SCAR-marker BC420, and subsequently tested these lines for CBB resistance in the greenhouse. Based on marker information, 28 of the 138 lines had the SCAR band present and were predicted to be resistant. Comparison of SCAR results with field data showed that 23 of 28 plants gave a resistant phenotypic reaction (rating<2.0), indicating an accuracy of 82%. Only 3.6% of the lines were misclassified as resistant. Cost estimates indicated that use of marker-assisted selections costs approximately one third less than greenhouse testing (Yu et al., 1999). Expression of QTL may differ over environments or populations in various crops (Park et al., 1999). Marker-QTL associations need to be confirmed in a breeding programme, particularly for traits such as CBB resistance that have complex inheritance patterns, low narrow-sense heritabilities and multiple genes involved (Park et al., 1999). Pyramiding of genes, expressing resistance to the same pathogen in a single cultivar, is necessary to achieve stable resistance. Use of marker-assisted selection can contribute considerably when pyramiding of genes is attempted (Dursun et al., 1995; Kelly and Miklas, 1999; Sing and Muñoz, 1999). Independence of resistance genes to be combined, however, needs to be closely monitored as many lines and cultivars have common sources of CBB resistance (Kelly and Miklas, 1999). The aim of this study was to determine whether SCAR-markers linked to four independent QTL derived from XAN 159 (SU91 and BC420) and GN #1 Nebr. sel 27 (SAP6 and BC409) could be used for indirect selection of resistance in the breeding programme in South Africa. MATERIALS AND METHODS Segregating populations of progenies from backcrosses were used for indirect selection of resistance using available SCAR markers SU91, BC420 (XAN 159 derived) and BC409 and SAP6 (GN # 1 Nebr. sel. 27 derived) (Table 1 and Table 3). Teebus and Kranskop were included as susceptible checks. Genotypes XAN 159, GN #1 Nebr. sel. 27, Wilk 2, Wilk 4, Wilk 6, Vax 3, Vax 4, Vax 5 and Vax 6 were used as resistant checks. A resistant line (48.15), developed in South Africa through interspecific crosses between P. vulgaris and P. acutifolius, as well as segregating populations with combined rust and CBB resistance (U 12 and C 18) were also included (Table 3). Phenotypic screening. Ten seeds of each line were planted in 20 litre plastic bags (4 seeds per bag) in sterile soil and maintained in a greenhouse at 18 oC night/28 oC day. Inoculum was prepared by suspending 48- to 72-hr-old cultures (isolates X6 and Xf105) in sterile distilled water, and adjusting it to 108 CFU ml-1 using a Shimadzu UV260 spectrophotometer. Fourteen to 20-day-old plants with fully expanded first trifoliate leaves were inoculated using the multiple needle inoculation method (Andrus, 1948). Inoculated plants were kept in a greenhouse at 18 oC night/28 oC day. Plants were rated for infection 14 days after inoculation on a 1 to 9 scale (Aggour et al., 1989) with 1 being resistant and 9 susceptible. Isolation of genomic plant DNA. Young leaves from resistant plants and susceptible checks were harvested and washed with sterile distilled water. Washed leaves were freeze-dried separately for each plant and kept at -20 oC until further use. DNA was isolated from sampled leaves using a modified version of the method described by Graham et al. (1994). Freeze-dried leaves were ground to fine powder for DNA extraction and a volume of 750 ml CTAB buffer [100 mM Tris, tris (hydroxymethyl) aminomethane], pH=8.0; 20 mM EDTA (ethylenediaminetetraacetate), pH=8.0; 1.4 mM NaCl; 2% (w/v) CTAB (hexadecyltrimethylammonium bromide); 0.2% (v/v) ß-mercaptho-ethanol added to approximately 250 ml of the fine leaf powder in a 1.5 ml microfuge tube. The suspension was thoroughly mixed and incubated at 65 oC for 1 hr. A 500 ml volume of chloroform: isoamyl alcohol (24:1) was added and the suspension mixed by gentle inversion. After centrifugation at 14 000 rpm for 3 min, the upper aqueous layer was transferred to a fresh tube containing 500 ml isopropanol, mixed by gentle inversion and incubated at room temperature for 20 min. The suspension was centrifuged at 14 000 rpm for 5 min, 500 ml 70% (v/v) ethanol added and incubated at room temperature for 20 min. DNA was precipitated at 14 000 rpm for 5 min, the pellet air-dried for 1 hr, and resuspended in TE buffer (10 mM Tris-Cl, pH 8.0; 1 mM EDTA, pH 8.0). Resuspended DNA was extracted with 1/10 volume 7.5 M ammonium acetate and an equal volume of chloroform: isoamyl alcohol (24:1). The aqueous layer was transferred to a fresh tube containing two volumes of cold absolute ethanol. Precipitated DNA was washed three times in cold 70% (v/v) ethanol, the pellet air-dried, and resuspended in TE buffer. DNA was treated with RNase for 2 hr at 37oC, after which concentration and purity were estimated by measuring absorbances at A260 and A280. DNA samples were diluted to a working solution of 200 ng ml-1. SCAR markers and PCR reactions. SCAR primers, BC409, SAP6, SU91 and BC420 (Table 2), were synthesised by GibcoBRL (Life Technologies, Glasgow, UK), based on primer sequences obtained from Miklas et al. (2000). Primers were suspended in TE buffer to a concentration of 200 pmol ml-1. A work solution of 10 pmol ml-1 was prepared. SCAR markers were used for the polymerase chain reaction (PCR) based on the protocol of Williams et al. (1990) with minor modifications. Amplification reactions were performed in a 25 ml reaction volume containing Promega (Promega Corporation, Madison, Wisconsin, USA) reaction buffer [500 mM KCl; 100 mM Tris-HCl, pH 9.0; 1% (v/v) Triton X-100], 2 mM MgCl2,100 mM of each dNTP (dATP, dCTP, dGTP and dTTP), 10 pmol primer, 1 unit Taq DNA polymerase (Promega) and 25 ng template DNA. Reactions were performed using a PCR Sprint Thermal Cycler (Hybaid Limited, UK) programmed for 5 min at 94 oC, 30 cycles of 1 min at 94 oC, 1 min at 58 oC for SAP6, SU91 and BC420 primers and 50 oC for BC409 primers, and 1.5 min at 72 oC, followed by one cycle of 5 min at 72 oC. Amplification products were analysed by electrophoresis in 1.5% (w/v) agarose gels (Seakem LE) at 80 V for 2 hr using UNTAN buffer (0.4 M Trisbase, 0.02 M EDTA, pH 7.4) and detected by staining with 1 mg ml-1 ethidium bromide. Gels were photographed under UV light with Polaroid 667 film. RESULTS Phenotypic disease reaction. Phenotypic reactions of lines are shown in Table 1. Genotypes XAN 159, Wilk 2, Wilk 4, Wilk 6, Vax 3, Vax 4, Vax 5 and Vax 6 were resistant to local Xap and Xapf isolates (ratings ranging from 1-2.7). GN #1 Nebr. sel. 27, Teebus and Kranskop were susceptible with mean disease ratings of 8.0, 9.0 and 7.5, respectively. Lines developed through backcross breeding were segregating and produced both susceptible and resistant plants. Susceptible plants were discarded and resistant plants used for molecular studies. Small white breeding lines (TCBR1, TCBR2, U12 and C18) were highly resistant (rating = 1.0). Large seeded red speckled sugar beans (PC 1470 BC2 1 and PC 1470 BC2 3.1) were moderately susceptible (rating = 4.3-4.9), while 48.1 and PC 2536-BC1 were resistant. Indirect screening. SCAR markers SAP6 and BC409 (GN #1 Nebr. sel. 27) were present in all the lines tested except for XAN 159, Wilk 2 and Wilk 4. Marker BC409 was absent in the breeding line PC 1470 BC2 3.1, while SAP6 was present in this line (Table 3). Marker SU91 (XAN 159 derived) was present in all the lines except Teebus, Kranskop, GN #1 Nebr. sel. 27 and 48.15 (Table 3). The other XAN 159 derived marker (BC420) was only present in XAN 159, Wilk 2, Wilk 4, Wilk 6, TCBR 1, TCBR 2, U12 and C18 (Table 3). DISCUSSION Results of this study indicate that existing markers for indirect selection of CBB resistance can be successfully used in the South African breeding programme. Presence of markers SAP6 and BC409 (GN #1 Nebr. Sel.27), in local cultivars Teebus and Kranskop, was most likely inherited from parents used to develop these cultivars. GN #1 Nebr. sel. 27 was derived from interspecific crosses between P. vulgaris and P. acutifolius and has been used in many breeding programmes as a resistance source (Coyne and Schuster, 1974; Mohan and Mohan, 1983). Although susceptible in South Africa, GN #1 Nebr. sel 27 and lines derived from it, have resistance in the USA (Coyne and Schuster, 1974) and Spain (C. Assensio, MBG-CSIC: personal communication). The Wilk lines were developed at Cornell University in USA and although the exact pedigree and germplasm used is not known, combined resistance genes from P. vulgaris, P. coccineus and P. acutifolius, including XAN 159 or its sister lines, were used in developing these lines (Singh and Muñoz, 1999). Presence of markers SU91 and BC420 in Wilk lines confirms that resistance in XAN 159 (or the same source) was used in developing these lines. Resistance in these lines was superior to that of XAN 159 when inoculated with local isolates, thus confirming the presence of additional CBB resistance genes. Presence of four markers, SU91, BC420, SAP6 and BC409, in small white canning bean lines (TCBR1, TCBR2, U12 and C18), developed through backcross breeding with XAN 159, confirmed successful transfer of resistance. Greenhouse results indicated that these lines had higher levels of resistance than XAN 159 (Table 1). This could be attributed to the combined resistance from GN Nebr. #1 sel. 27 and XAN 159 present in these lines. Although GN #1 Nebr. sel. 27 is susceptible to local isolates when tested on its own, it seems to contribute to higher levels of resistance when combined with XAN 159 resistance. Similar results were obtained by Mutlu et al. (2005). Studies conducted by O’Boyle and Kelly (2007), however, indicated that presence of both SU91 and BC420 markers resulted in lower levels of CBB resistance than provided by either marker alone. Marker BC420 was absent in XAN 159 derived Kranskop-lines (PC 1470 BC2 1 and PC 1470 BC2 3.1) with acceptable seed colour (red speckled seed). These lines were only moderately resistant when tested in the greenhouse, indicating that the QTL linked to this marker is important in order to obtain high levels of resistance. Resistance of Kranskop lines, however, has been improved and this can be attributed to presence of the QTL linked to the SU91 marker. The BC420 marker is located near the V-locus conditioning purple flower colour (Miklas et al., 2000). High levels of resistance have been identified in some Kranskop derived lines (Fourie, unpublished data) but highly resistant plants had purple flowers resulting in plants producing seed with unacceptable colour. Mutlu et al. (2005) experienced similar problems in obtaining acceptable seed coat colour in pinto bean lines. The BC420 marker was also absent in the Vax lines. These lines were developed at the International Center for Tropical Agriculture (CIAT) and are highly resistant to Xap and Xapf (Singh and Muñoz, 1999). Absence of marker BC420 could indicate that the linkage between the V-locus and the resistance gene has been broken and that although the resistant gene may be present, the marker is absent. The possibility also exists that other untagged genes could contribute to the high levels of resistance present in these lines, as resistant genes from different sources have been pyramided into these lines (Singh and Muñoz, 1999). Resistance from Vax lines should be used to improve resistance in large seeded (red speckled sugar) bean varieties. Progenies from backcrosses with Kranskop as recurrent parent using Vax 4 exhibited high levels of resistance when tested in the greenhouse and presence of all markers found in Vax 4 confirms transfer of resistance. The locally developed line 48.15, developed through interspecific crosses between P. vulgaris and P. acutifolius, was highly resistant when tested in the greenhouse. PCR studies indicated that resistance was not the same as XAN 159 (markers absent) and attempts should be made to combine this resistance in XAN 159 derived Kranskop lines. XAN 159 derived CBB resistant Teebus lines have been successfully combined with rust resistant Teebus lines developed in an independent breeding programme. Markers are also available to confirm rust resistance (Stavely, 2000). The use of markers is especially advantageous when combining resistance to different diseases into one cultivar. Results gained from this study show that marker-assisted selection can be implemented in breeding for common bacterial blight resistance in South Africa. The use of molecular markers alone, however, has not resulted in lines with resistance superior to that of XAN 159. The combined use of both phenotypic screening and Molecular markers in breeding for bean common bacterial blight resistance molecular markers is, therefore, important in developing CBB resistant lines. ACKNOWLEDGEMENT Partial support for this work provided by the Pan-Africa Bean Research Alliance (PABRA) is highly appreciated. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11034t3.jpg] [cs11034t2.jpg] [cs11034t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}