 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
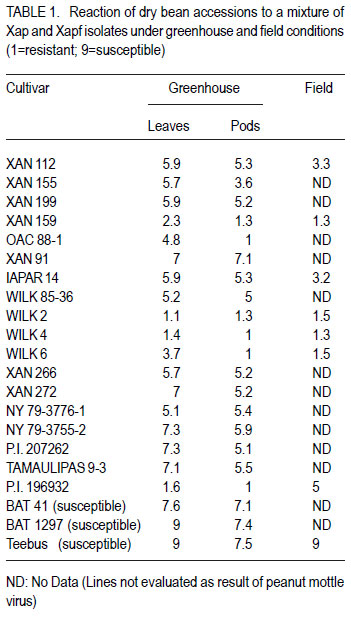
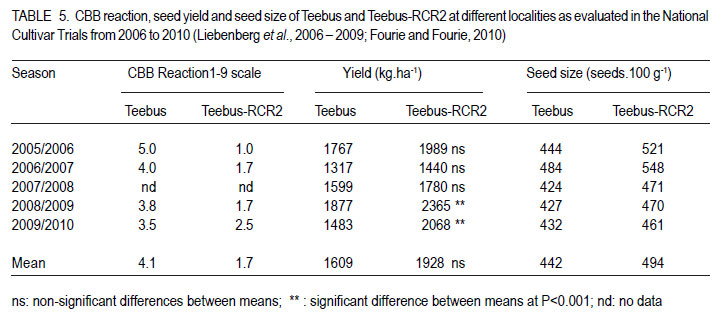
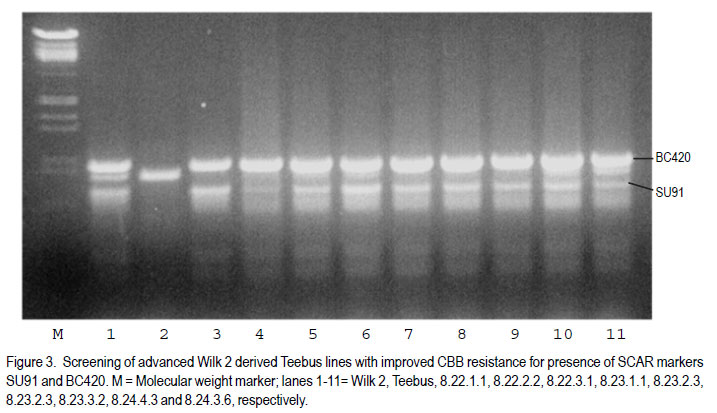
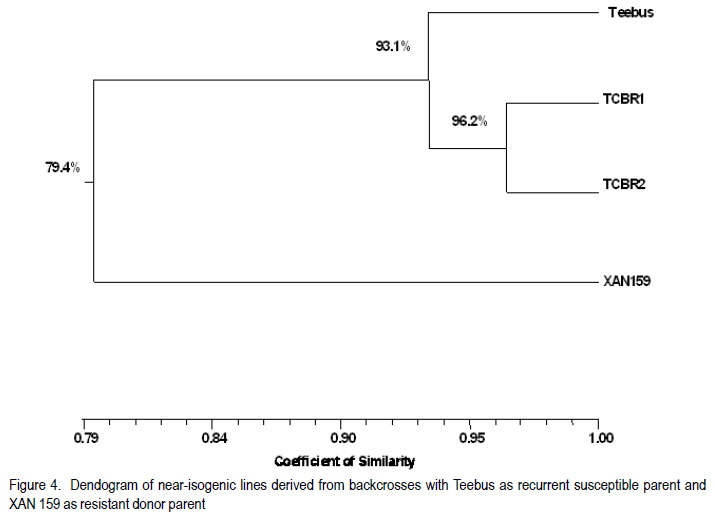
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 377-386 Improvement of common bacterial blight resistance in South African dry bean cultivar teebus D. Fourie, L. Herselman1 and C. Mienie Agricultural Research Council-Grain Crops Institute, Private Bag X1251, Potchefstroom, 2520, South Africa Code Number: cs11035 ABSTRACT Common bacterial blight (CBB) caused by Xanthomonas axonopodis pv. phaseoli is an important seed-borne disease of dry beans in South Africa. Development of resistant cultivars is considered the best control measurement for the disease. Backcross breeding was used to improve CBB resistance in the small white canning bean, cv. Teebus, using resistance sources XAN 159 and Wilk 2. High resistance levels in near-isogenic lines, developed in two independent breeding programmes, indicated successful transfer of resistance from both sources. Presence of SCAR-markers, SU91 and BC420, in 35 of 39 XAN 159 derived Teebus lines and all lines derived from Wilk 2, confirmed successful resistance transfer. AFLP studies conducted to determine genetic relatedness of two near-isogenic Teebus lines, showed a similarity of 96.2% with the maximum similarity between these lines and Teebus being 93.1%. One cultivar, Teebus-RCR2 with yield similar to Teebus and improved resistance to CBB has been released from the programme. Key Words: Backcross breeding, Phaseolus vulgaris, Xanthomonas axonopodis RÉSUMÉ Le flétrissement bactérien commun (CBB) causé par Xanthomonas axonopodis pv. phaseoli est une importante maladie de grains des haricots secs en Afrique du Sud. Le développement des cultivars résistants est considéré comme une meilleure mesure de contrôle de la maladie. L’amélioration par croisement en retour était utilisée pour améliorer la résistance au CBB dans le petit haricot blanc, cv.Teebus, en utilisant des sources résistantes XAN 159 et Wilk 2. De niveaux élevés de résistance dans des lignées proches isogéniques, développées dans deux programmes indépendants d’amélioration, ont indiqué un transfert réussi de résistance de toutes ces deux sources. De la présence des marquers SCAR, SU91 et BC 420 dans 35 de 39 XAN 159 a découlé les lignées Teebus et toutes les lignées dérivant de Wilk 2, confirmant ainsi un transfert réussi de résistance. Des études AFLP conduites pour déterminer la relation génétique de deux lignées proches isogéniques Teebus, ont montré une similarité de 96.2% avec la maximum de similarité entre ces lignées et Teebus de 93.1%. Un cultivar, TeebusRCR2 avec rendement similaire au Teebus et résistance améliorée au CBB a été disseminée par le programme. Mots Clés: Amélioration par croisement en retour, Phaseolus vulgaris, Xanthomonas axonopodis INTRODUCTION One of the most important dry bean (Phaseolus vulgaris L.) diseases in South Africa is common bacterial blight (CBB), caused by the bacterium Xanthomonas axonopodis pv. phaseoli (Xap) (Smith) Vauterin, Hoste, Kosters; and Swings and its fuscans variant, X. axonopodis pv. phaseoli var. fuscans (Xapf) (Fourie, 2002). The disease is widespread worldwide and occurs in all the major South African bean producing areas (Fourie, 2002). Yield losses have been poorly documented but are reported to vary between 22 and 45% (Wallen and Jackson, 1975; Yoshii, 1980). Infected seed is the primary inoculum source and planting of pathogen-free seed is an important means of disease avoidance. The most effective and economic CBB control strategy is the use of genetic resistance (Miklas et al., 2003; Singh and Muñoz, 1999). CBB resistance breeding has been extensively researched (Beebe and Pastor-Corrales, 1991). Rands and Brotherton (1925) first identified lines with CBB resistance. Subsequent efforts yielded moderate levels of resistance (Yoshii et al., 1978) with no immunity in P. vulgaris. Higher levels of resistance were found in scarlet runner bean (P. coccineus), with the highest levels identified in tepary beans (P. acutifolius) (Singh and Muñoz, 1999). Interspecific crosses between P. vulgaris and P. acutifolius resulted in development of resistant lines such as GN #1 Nebr. sel. 27, XAN 112, XAN 159, XAN 160, XAN 161 and OAC 88-1 (Schuster and Coyne 1981; Silva et al., 1989; Beebe and Pastor-Corrales, 1991). Varieties with resistance were also developed from interspecific crosses between P. vulgaris and P. coccineus (Park and Dhanvantari, 1987; Miklas et al., 1994). Resistant lines (Vax 1, Vax 2, Vax 3, Vax 4, Vax 5 and Vax 6) were developed at the International Centre for Agriculture in the Tropics (CIAT) from interspecific hybridisation of P. vulgaris and P. acutifolius and gene pyramiding (Singh and Muñoz, 1999). These lines showed high levels of resistance when tested against isolates from various geographical origins (Zapata et al., 1998; Jara et al., 1999). Vax 1 and Vax 2 were, however, susceptible when evaluated in Uganda (R. Buruchara, CIAT: personal communication) and South Africa (D. Fourie: unpublished data). Resistance levels in Vax 3, Vax 4 and Vax 6 are as high as those found in P. acutifolius (Singh and Muñoz, 1999). Most of the resistant sources are considered exotic germplasm and are poorly adapted to local conditions, but are suitable as donor parents in a breeding programme. Depending on resistance source and evaluation methodology used, one to six genes appear to confer CBB resistance in bean ( Adams et al., 1988; Silva et al., 1989; Eskridge and Coyne, 1996). Genetic markers have indicated that CBB resistance is linked to between two and six quantitative trait loci (QTL) (Jung et al., 1996; Park et al., 1998; Tsai et al., 1998). CBB resistance is quantitatively inherited with dominance for susceptibility (Finke et al., 1986). Although gene action is primarily additive, dominance and epistatic effects have been observed (Beebe and Pastor-Corrales, 1991). Low estimates of narrow sense heritability have also been reported (Arnaud-Santana et al., 1994). All locally grown commercial dry bean cultivars are susceptible to CBB (Fourie, 2011) and improvement of resistance in local cultivars is important for the control of CBB. Thus, the aim of this study was to identify sources of CBB resistance in exotic germplasm for use in a backcross breeding programme to improve resistance of local commercial varieties. MATERIALS AND METHODS Evaluation of germplasm for CBB resistance. Eighteen CBB resistance sources (Table 1), obtained from CIAT were screened, under field and greenhouse conditions for resistance to local isolates of Xap and Xapf. BAT 41 and BAT 1297, obtained from CIAT and a South African cultivar, Teebus was included as susceptible checks. Greenhouse screening. Twenty-five seeds of each genotype were planted in 20 litre plastic bags (5 seeds per bag) in sterile soil and maintained in a greenhouse at 18 °C night/28 °C day. Seedlings were thinned to four plants per pot after emergence. A mixture of two local aggressive isolates (X6 and Xf105) was used for inoculation. Inoculum was prepared by suspending 48 to 72hr-old cultures in sterile distilled water, which was adjusted to 108 CFU ml-1. Fourteen to 20-day-old plants with fully expanded first trifoliate leaves were inoculated using the multiple-needle inoculation method (Andrus, 1948). Control plants were inoculated with sterile distilled water. Plants were maintained in a greenhouse at 18 °C night/28 °C day and rated, on a 1 to 9 scale (Aggour et al., 1989), 14 days after inoculation, with 1 being resistant and 9 susceptible. Young, detached pods from each genotype were inoculated with one Xap isolate (X6) using the method of Aggour et al. (1989). Disease reactions were recorded 10 days after inoculation on a 1-9 scale (Aggour et al., 1989) with 1 being resistant and 9 susceptible. Field screening. Two 5-m rows (65 seeds per row) of each genotype were planted in an unreplicated trial in the field and evaluated for CBB resistance. Inoculum was prepared similar to that for the greenhouse trials with the exception that non-sterile tap water was used. A motorised backpack sprayer was used for inoculating plants in the field at 25, 32 and 39 days after planting. Rows were evaluated for disease reaction from the time when first symptoms appeared until the crop matured. Evaluations were based on the CIAT 1-9 scale with 1 being resistant and 9 susceptible (Van Schoonhoven and Pastor-Corrales, 1987). Breeding for resistance. Genotypes exhibiting highest levels of resistance to local isolates under greenhouse and field conditions were selected to improve resistance of a local cultivar, Teebus, (small white canning bean) (Table 2). Teebus was selected based on its commercial value and preference by the canning industry (Liebenberg et al., 1999) Crosses were made in the greenhouse between the resistant donor (pollen) parent and the recurrent susceptible parent (Teebus). First trifoliate leaves of plants from F1-generations were inoculated with a bacterial suspension containing approximately 108 CFU ml-1 water, using the multiple needle puncture method. Leaves were rated for infection 14 days after inoculation on a 1 to 9 scale, with 1 being highly resistant and 9 being highly susceptible. Teebus plants were inoculated as susceptible controls. Susceptible plants were discarded (plants rated >3-9) and resistant plants (rated 1-3) retained for backcrossing. Backcrossing to the recurrent parent was continued for five generations and approximately 94% of the recurrent parent’s genome was recovered with addition of the resistance gene(s). Segregating BC5F2 populations were planted in field trials at Potchefstroom in South Africa during the 1999/2000 season and evaluated for resistance. Plots consisted of unreplicated single rows of 5 m each with 30 seeds planted per row. Teebus was planted every sixth row throughout the plot as a susceptible check. First or second trifoliate leaves of each plant in a 5 m row were inoculated using the multiple needle method, which was followed by spray inoculating plants with a motorized backpack sprayer. Spray inoculations were repeated weekly until adequate disease developed on susceptible checks. Each plant was rated separately and single plants with high levels of resistance (rating 1-2) were marked. Spray-inoculated canopies of selected single plants were evaluated periodically from when first symptoms appeared on the susceptible checks until the crop matured. Single plant progeny rows (F3 generation) were planted during winter (May, 2000) at Makhatini Research Station, KwaZulu-Natal in South Africa, inoculated and similarly rated. Single plants were again selected and F4 generations planted in progeny rows at Potchefstroom the following summer (2000/2001). A total of 1972 single plant field selections were made from advanced Teebus lines (BC5F2-F5) with resistance from XAN 159. Six hundred and forty three single plant progeny rows were evaluated and 136 lines (BC5F6) lines judged to be homozygous and with high levels of resistance (rating 1-2) were evaluated for yield in checkrow trials. A total of 401 single plants were selected from Wilk 2 derived Teebus lines (BC5F2-F5), from which 146 single plant progeny rows were evaluated and 11 homozygous resistant lines selected for further evaluation. Crosses were made between Teebus lines with improved resistance to CBB and Teebus lines with improved resistance to rust developed in a separate breeding programme (data not shown). Progenies from these crosses were extensively screened for resistance in greenhouse and field trials, and from these 79 high yielding lines with acceptable levels of resistance to CBB and rust were entered into advanced yield trials during the 2003/04 season and from these 17 lines were selected for further evaluation during the 2004/05 season. The best performing variety selected from the advanced yield trials was entered in the National Cultivar Trials and evaluated over five seasons in multiple locations (between 18 and 28 locations – varied annually). Confirmation of resistance using SCAR markers. Thirty nine near-isogenic resistant Teebus lines (BC5F4), derived from backcrossing with XAN 159 as donor parent, and eight lines derived from backcrossing with Wilk 2 (BC5F2), were evaluated for presence of two independent CBB resistant QTL (SCAR markers SU91 and BC420) (Miklas et al., 2000) to confirm transfer of resistance in early generations (Table 3). The methods used were similar to that described by Fourie and Herselman (2011). Genetic relatedness of near-isogenic Teebus lines. Extracted DNA from Teebus, XAN 159 and two near-isogenic (BC5F6) high yieldingTeebus lines (TCBR1 and TCBR2) with improved CBB resistance were subjected to amplified fragment length polymorphism (AFLP) analysis to determine genetic distances between these lines. AFLP adapters and primers (Table 4) were designed based on the methods of Vos et al. (1995). Primers were synthesized by GibcoBRL (Life Technologies, Glasgow, UK) and oligonucleotides used for adapters were PAGE (polyacrylamide gel electrophoresis) purified. Adapters were prepared by adding equimolar amounts of both strands, heated for 10 min to 65°C in a water bath and left to cool at room temperature. Gel electrophoresis for AFLP analysis was performed (Vos et al., 1995) using a 5% (w/v) denaturing polyacrylamide gel [19:1 acrylamide: bis-acrylamide; 7 M urea; 1x TBE buffer (89 mM Tris-borate; 2.5 mM EDTA)]. Electrophoresis was carried out at constant power, 80 W for approximately 2 hr. Polyacrylamide gels were silver-stained following the protocol described by Silver SequenceTM DNA Sequencing System manual supplied by Promega (Madison, WI, USA). Gels were left upright overnight to air-dry and photographed by exposing photographic paper (Kodak Polymax II RC) directly under the gel to about 20 sec of dim light. This produced a negative image, exactly the same size as the gel. AFLP data were scored based on presence (1) or absence (0) of DNA fragments obtained for each line. Only reliable and repeatable fragments were considered. Pair-wise genetic distances were calculated between isolates (Nei and Li, 1979). Cluster analysis was done using the unweighted paired group method using arithmetic averages (UPGMA). Statistical analyses of AFLP data were performed using NTSYSpc version 2.02i. RESULTS Germplasm evaluation for CBB resistance. The reaction of genotypes to local Xap and Xapf isolates is shown in Table 1. Four lines, XAN 159, Wilk 2, Wilk 4 and Wilk 6 exhibited good combined leaf, pod and field resistance. P.I.196932 was resistant when tested in the greenhouse, but was moderately susceptible in the field. The susceptible checks (BAT 41, BAT 1297 and Teebus) were truly susceptible under both greenhouse and field conditions. Thirteen lines were lost due to peanut mottle virus (Table 1) and could, therefore, not be evaluated in the field for CBB resistance. Two lines, XAN 155 and OAC 88-1 were moderately susceptible when inoculated on first trifoliate leaves but were moderately to highly resistant when inoculated on pods. Breeding for resistance. XAN 159 and Wilk 2 were selected for their high levels of resistance to local isolates (Table 1), for use in two independent backcross programmes to improve resistance of cv. Teebus. Five backcrosses were completed in both breeding programmes and approximately 93% of Teebus has been recovered with improved CBB resistance. Crosses between the CBB resistant lines and rust resistant lines developed in a separate breeding programmes were successfully made and progenies with combined CBB and rust resistance selected (data not shown) for evaluation in advanced yield trials. One cultivar, Teebus-RCR2 (Reg. No. ZA 20053277) was released in April 2005. Data from the National Cultivar Trials conducted over multiple years and locations are shown in Table 5. Common bacterial blight resistance in TeebusRCR2 (1.7) was superior to that of Teebus (4.1) when rated on a 1-9 scale, with 1 being resistant and 9 being susceptible. Teebus-RCR2 was higher yielding than Teebus, however, significant differences in yield were only obtained during the 2008/2009 and 2009/2010 seasons (Table 5). Confirmation of resistance using SCAR markers. SCAR-marker SU91 was present in all (39) XAN 159 derived Teebus lines tested, while BC420 was present in 35 of 39 lines (Fig. 1 ). Both SU91 and BC420 markers were present in all (eight) lines derived from Wilk 2 backcrosses with Teebus as recurrent parent (Fig. 2). Genetic relatedness of near-isogenic Teebus lines. The dendrogram (Fig. 3), drawn from AFLP data, resulted in two groups; one containing the resistant donor parent (XAN 159) and the other containing near-isogenic lines and the recurrent susceptible parent (Teebus). The Teebus cluster was linked to the XAN 159 cluster at a similarity of 79.4%. Near-isogenic lines (TCBR1 and TCBR2) exhibited a similarity of 96.2%. Similarity between the two near isogenic lines and Teebus was 93.1% suggesting a good fit between the dendrogram and the genetic similarity matrices. DISCUSSION Adequate levels of resistance were identified in XAN 159 and Wilk 2 (Table 1) for use in a backcross breeding programme to improve resistance of cv. Teebus. XAN 159 was developed at CIAT through interspecific crosses between P. vulgaris and P. acutifolius, which exhibited combined leaf and pod resistance to local isolates. Similar resistance in XAN 159 was obtained by Arnaud-Sanata et al. (1993), when evaluating 18 lines for combined leaf and pod resistance in the USA. Resistance instabilities have been reported in XAN 159 and its progenies (Beebe and Pastor-Corrales, 1991), however, it is still used widely in resistance breeding programmes (Beebe and Pastor-Corrales, 1991; Park et al., 1998; Mutlu et al., 1999; Singh and Muñoz, 1999). Wilk 2 has combined resistance genes from P. vulgaris, P. coccineus and P. acutifolius, including XAN 159 or its sister lines (Singh and Muñoz, 1999) and was developed at Cornell University, USA. Differential reactions of pods and leaves in a number of genotypes screened (Table 1) indicated the importance of evaluating both these plant parts when developing resistant plants. Similar differential reactions of pods and leaves to Xap have been reported previously (Schuster et al., 1983; Aggour et al., 1989). Phenotypic disease reaction of advanced lines (rating 1-2), in greenhouse and field evaluations, confirmed transfer of resistance from both XAN 159 and Wilk 2 and indicated that resistance in cv. Teebus was successfully improved. Homozygous resistant lines were selected and an improved Teebus variety (Teebus-RCR2) with resistance to CBB and rust was released. TeebusRCR2 was higher yielding than Teebus and although significant differences were only observed during two seasons, the mean increase in yield over the five seasons were approximately 300 kg ha-1. This is advantageous to farmers especially to those producing beans in areas where CBB is problematic. PCR studies indicated that both existing SCAR-markers, SU91 and BC420, were present in XAN 159 and Wilk 2 derived Teebus lines tested. This confirms successful transfer of resistance in these advanced lines. Greenhouse results indicated that these lines had resistance levels superior to that of XAN 159 (data not shown), which could be attributed to the presence of additional resistance gene(s) being present in these lines. Presence of XAN 159 markers in Wilk 2 derived lines confirms that XAN 159 or similar source was used in developing Wilk 2. Since Wilk lines were the first with pyramided resistance genes from various sources, additional CBB resistance genes might be present in advanced Teebus lines. A combination of XAN 159 and Wilk 2 derived Teebus lines may result in stable CBB resistance. However, markers linked to additional resistance genes in Wilk 2 are necessary when gene pyramiding is attempted. High genetic relatedness between Teebus and near-isogenic lines as indicated in AFLP studies indicated that characteristics of cv. Teebus have been recovered with the addition of the resistance gene(s) from XAN 159. Breeding for resistance in canning beans, however, should always progress within the boundaries set by the industry for canning quality. It is, therefore, important to maintain, as far as possible, the sought-after quality of the original cultivar. Although Teebus-RCR2 was acceptable for canning purposes in small scale canning tests, it failed in certain critical factory tests (Liebenberg et al., 2009; Fourie and Fourie, 2010). Negative correlation with regard to quality, has been reported where XAN 159 was used as resistance source (J.D. Kelly, Michigan State University: personal communication). ACKNOWLEDGEMENT Partial support for this work provided by the Pan-Africa Bean Research Alliance (PABRA) is highly appreciated. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11035t2.jpg] [cs11035f1.jpg] [cs11035f3.jpg] [cs11035f2.jpg] [cs11035t5.jpg] [cs11035t3.jpg] [cs11035t1.jpg] [cs11035t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}