 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
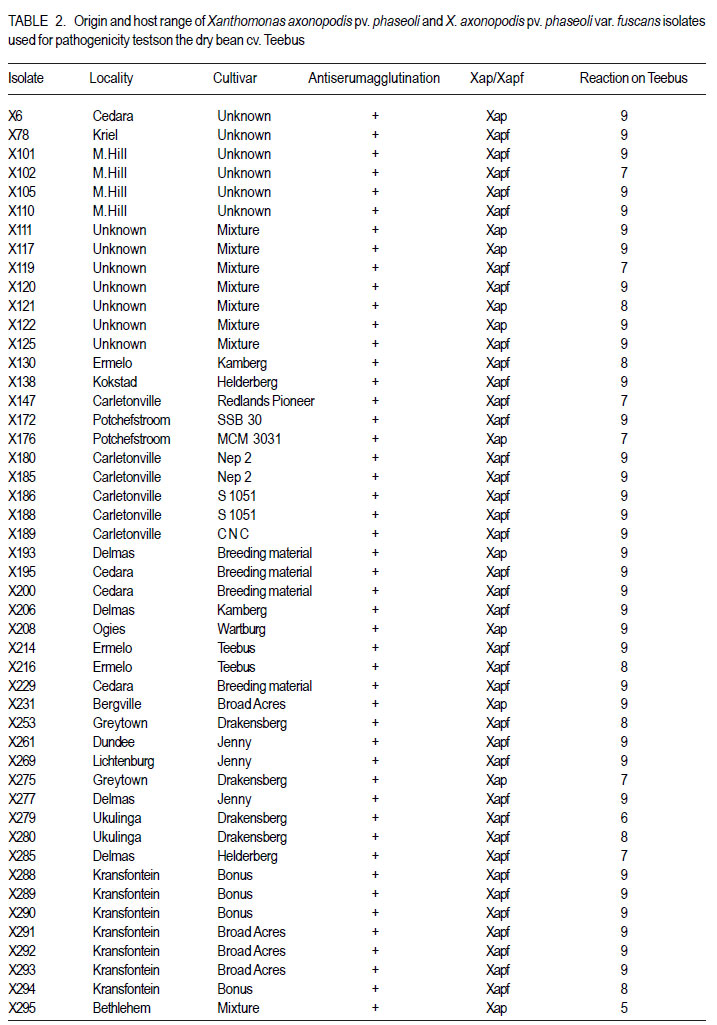
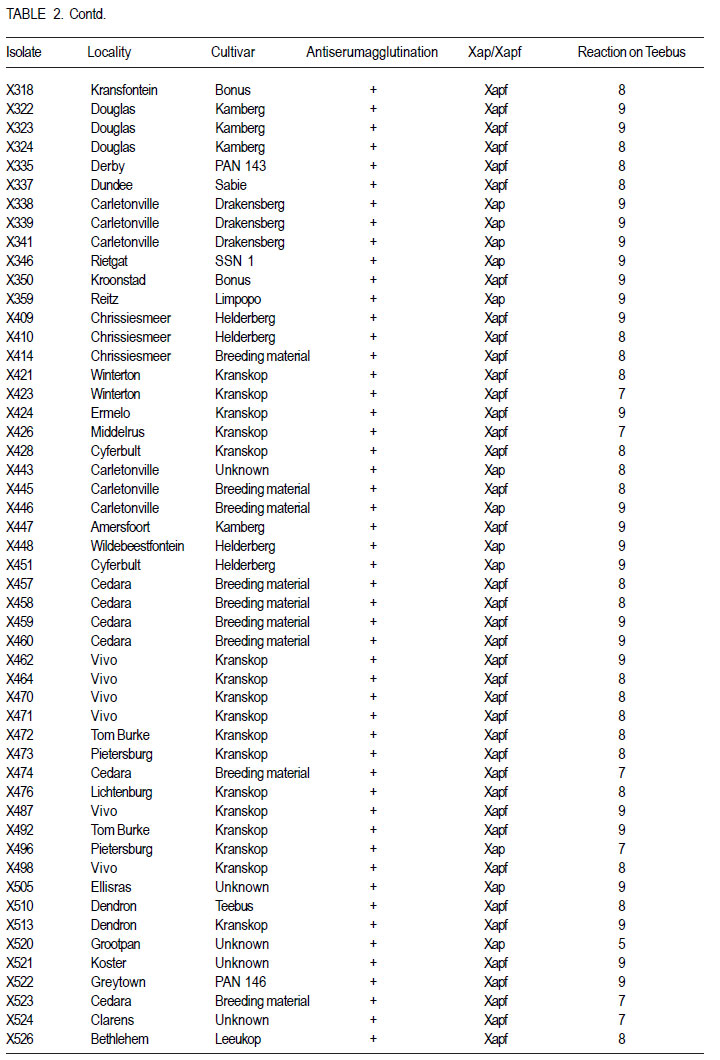
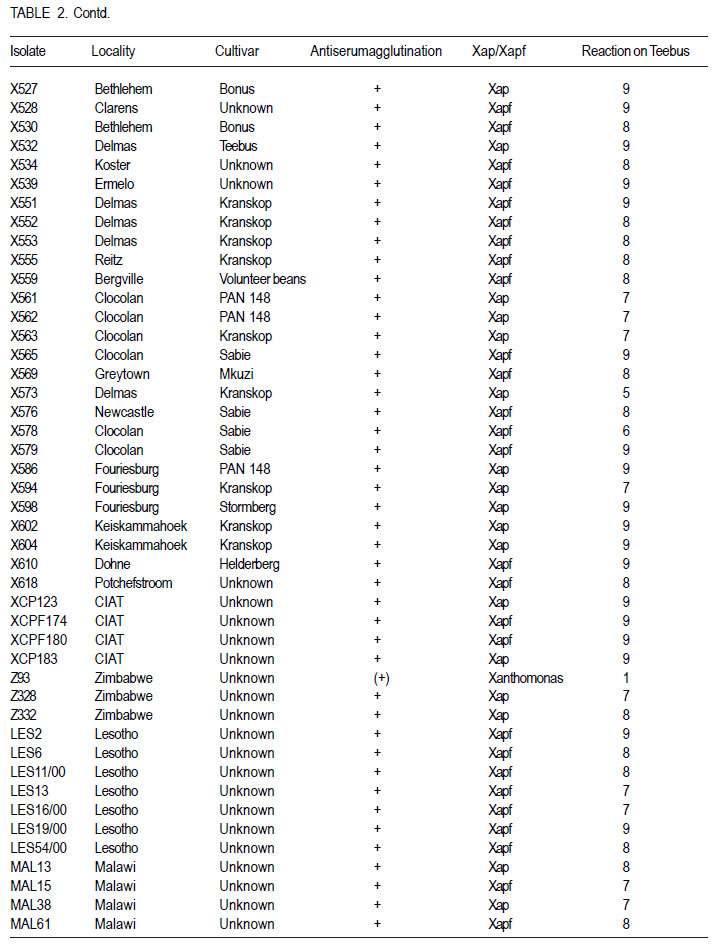
African Crop Science Journal, Vol. 19, No. 4, 2011, pp. 393-407 Pathogenic and genetic variation in Xanthomonas axonopodis pv. phaseoli and its fuscans variant in Southern Africa D. Fourie and L. Herselman1 Agricultural Research Council-Grain Crops Institute, Private Bag X1251, Potchefstroom, 2520, South Africa
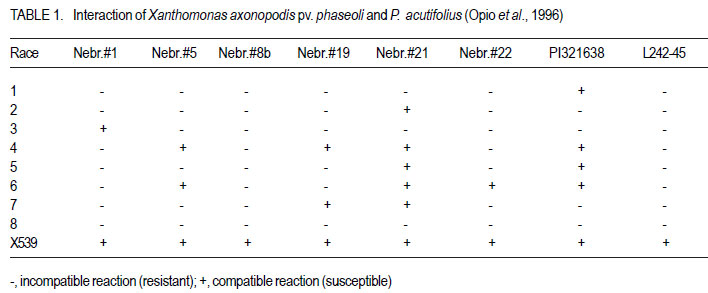
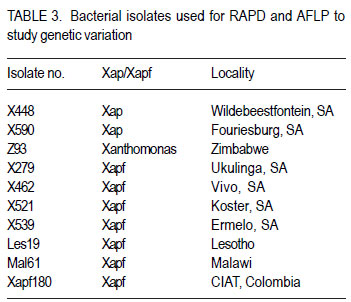
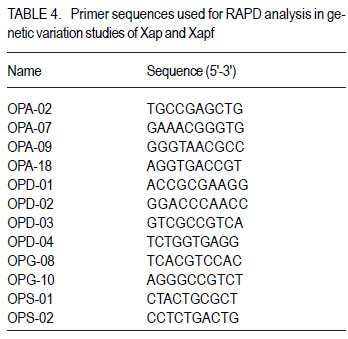
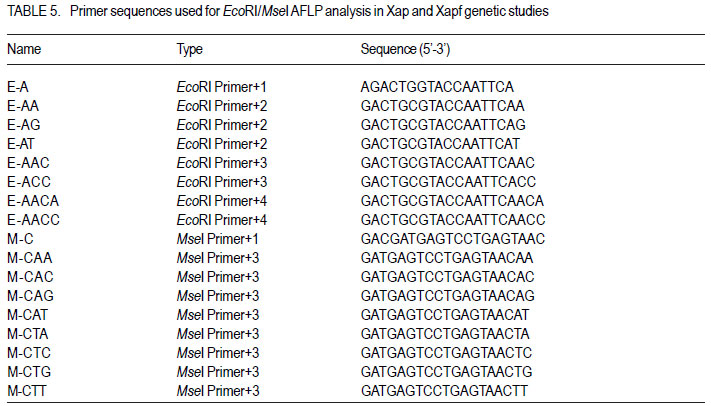
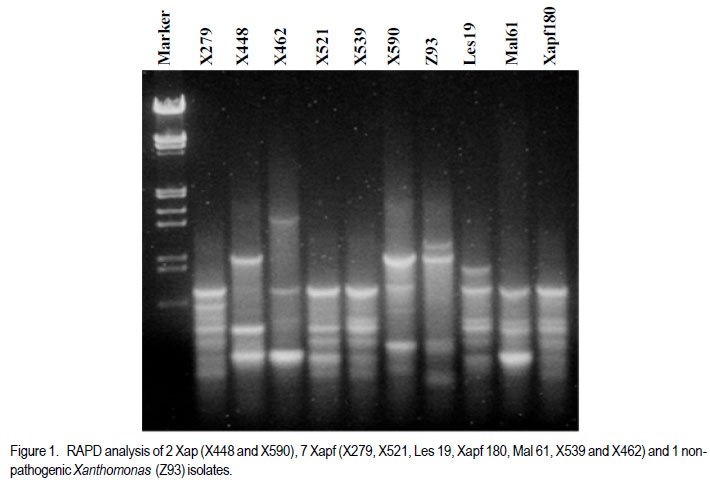
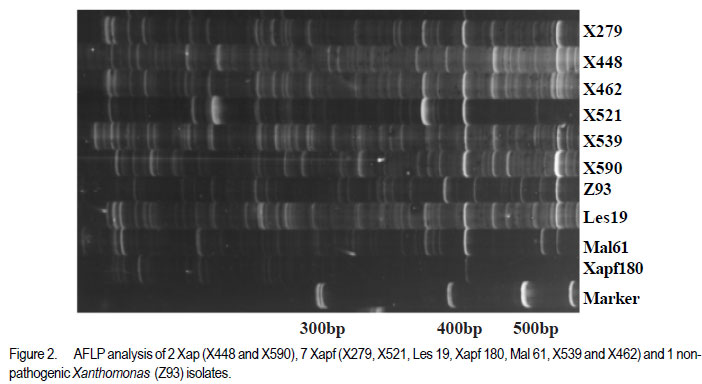
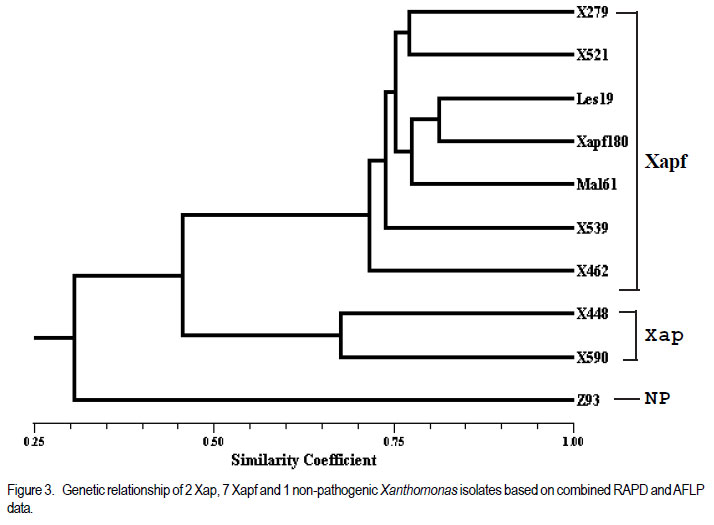
Code Number: cs11037 ABSTRACT Common bacterial blight (CBB) caused by Xanthomonas axonopodis pv. phaseoli and its fuscans variant, X. axonopodis pv. phaseoli var. fuscans is a widespread disease of dry beans in South Africa. Variation within pathogen populations has been reported and in order to breed for resistance it is important to investigate whether variation exists within the local pathogen population. One hundred and forty three common bacterial blight isolates from 44 localities in four countries, were inoculated onto eight Phaseolus acutifolius lines that differentiate between pathogenic races. This differential set was expanded to include resistant genotypes, XAN 159, GN #1 Nebr. sel 27, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6 and cv. Teebus as susceptible check. Genetic variation within nine selected Xap and Xapf isolates and a non-pathogenic Xanthomonas isolate, was studied using RAPD and AFLP analysis. Genotypes XAN 159, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6 were resistant to all isolates, while GN #1 Nebr. sel 27 and cv. Teebus were susceptible. Isolates varied in aggressiveness on cv. Teebus; however, the pathogenic reaction on the set of differentials indicated that all, but one isolate, grouped in what has been reported as race 2. Thus, results based on reaction of the majority isolates, suggest the absence of different races. However, the distinct differential reaction recorded for a single isolate, may prove to represent another, as yet unrecorded, race of this pathogen. Both RAPD and AFLP analyses revealed high frequency of DNA polymorphism among isolates and could distinguish between Xap, Xapf and a non-pathogenic isolate. Differences between Xap and Xapf isolates demonstrate that these are two distinct groups of bacteria. Key Words: AFLP analysis, common bacterial blight, P. acutifolius, Phaseolus vulgaris, RAPD analysis RÉSUMÉ Le flétrissement bactérien commun (CBB) causé par Xanthomonas axonopodis pv. Phaseoli et ses variantes “fuscans”, X. axonopodis var. pv. phaseoli est une maladie répandue des haricots en Afrique du Sud. Pour améliorer la résistance à la maladie, une étude était menée pour évaluer la variation à l’intérieur de populations pathogènes locales. Pour ce faire, cent quarante trois isolats de bactéries communes de flétrissement issues de 144 localités de quatre pays étaient inoculés dans huit lignées de Phaseolus acutifolius différentes de races pathogéniques. Cet ensemble differentiel était étendu afin d’inclure les génotypes résistants XAN 159, GN #1 Nebr. sel 27, Wilk 2, Wilk 6, Vax 4, Vax 5 et Vax 6 ainsi que cv. Teebus comme témoins susceptible. La variation génétique parmi neuf Xap et leurs isolats sélectionnés ainsi qu’un isolat de Xanthomonas non-pathogénique étaient étudiés par l’analyse RAPD et AFLP. Les génotypes XAN 159, Wilk 2, Wilk 6, Vax 4, Vax 5 et Vax 6 étaient résistants à tous les isolats alors que GN #1 Nebr. sel 27 et cv. Teebus étaient susceptibles. Les isolats variaient dans leur agressivité au cv; par ailleurs, la réaction pathogénique sur l’ensemble des différentiels a indiqué que tous les isolats sauf un seul, pouvaient être groupés dans ce qui était décrit comme race 2. Ainsi, les résultats basés sur la réaction de la majorité des isolats suggèrent l’absence de différentes races. Par ailleurs, la réaction différentielle distincte enregistrée dans un seul isolat, pourrait permettre de représenter une autre race de ce pathogène non encore enregistrée. Les analyses RAPD et AFLP ont révélé une fréquence élevée du polymorphisme de l’AND parmi les isolats et pourraient permettre de faire une distinction entre Xap, Xapf et un isolat non pathogénique. Les différences entre les isolats Xap et Xapf démontrent que ces derniers forment deux groupes distincts de bactéries. Mots Cles: Analyse AFLP, bactérie commune de flétrissement, P. acutifolius, Phaseolus vulgaris, analyse RAPD INTRODUCTION Common bacterial blight (CBB) caused by Xanthomonas axonopodis pv. phaseoli (Xap) (Smith) Vauterin, Hoste, Kosters and Swings and its fuscans variant, X. axonopodis pv. phaseoli var. fuscans (Xapf), is a devastating seed-borne disease of dry beans (Phaseolus vulgaris) in many parts of the world (CIAT, 1985). The disease is widespread throughout the South African production areas (Fourie, 2002) and is favoured by high temperatures and high relative humidity (Sutton and Wallen, 1970). In eastern and southern Africa, common blight has been reported in 19 of the 20 bean producing countries (Allen, 1995) and is considered one of five most important and widespread biotic constraints to dry bean production in sub-Saharan Africa (Gridley, 1994). Genetic resistance is considered the most effective and economical strategy for the control of bean common blight (Rands and Brotherton, 1925). However, deployment of resistance without knowledge of variation within a pathogen population could result in costly failure (Taylor et al., 1996). Pathogenic variation in Xap and Xapf isolates has been demonstrated in several reports (Schuster and Coyne, 1971; Schuster et al., 1973; Yoshii et al., 1978; Schuster, 1983; Jindal and Patel, 1984; Mkandawire et al., 2004; López et al., 2006; Mutlu et al., 2008). Ekpo and Saettler (1976) indicated that Xapf isolates were more pathogenic than Xap. These differences in pathogenicity have been confirmed by other investigators (Leakey, 1973; Bozzano-Saguier and Rudolph, 1994; Opio et al., 1996; Mkandawire et al., 2004; Mutlu et al., 2008), but it has been suggested that the brown pigment is not associated with pathogenicity (Gilbertson et al., 1991; Tarigan and Rudolph, 1996) and should be considered of lesser pathological importance (Schuster and Coyne, 1975). Gilbertson et al. (1991) studied genetic diversity in isolates of Xap and Xapf, using DNA probes isolated from a single Xap isolate genome on isolates from different geographical locations. These studies indicated that there are two distinct groups of bacteria. However, similarities between isolates were revealed when probes were hybridised to DNA from other X. campestris pathovars, indicating sufficient similarity to consider Xapf a variety of Xap (Gilbertson et al., 1991). Reports of physiological specialisation in P. vulgaris have been contradictory. Zapata (1996) indicated P. vulgaris genotypes that are useful in differentiation of Xap. However, evidence exist suggesting quantitative interactions between Xap and P. vulgaris (Opio et al., 1996). Host specialisation of Xap reactions on tepary (P. acutifolius) lines has been reported (Zapata and Vidaver, 1987; Zaiter et al., 1989; Opio et al., 1996) with eight physiological races identified, suggesting a gene-for-gene relationship (Opio et al., 1996). Despite this gene-for-gene interaction, resistance to Xap and Xapf in P. vulgaris, derived from P. acutifolius, has remained non-specific and durable (Opio et al., 1996). Tepary bean is an excellent source of resistance due to high resistance levels to Xap and Xapf. Variation that may exist in the local pathogen population is important when selecting parents with resistance originating from tepary cultivars. The aim of the study was to determine pathogenic and genetic variation in Xap and Xapf isolates in southern Africa ensuring that appropriate resistance sources are deployed when developing CBB resistant cultivars. MATERIALS AND METHODS Isolation and identification of isolates. Diseased plant material (at different growth stages depending on the time the disease was noted) was collected from major bean production areas in South Africa; and Malawi, Lesotho and Zimbabwe during the 2000/2001 and 2001/2002 seasons (Table 2a, 2b, 2c). The infected material were rinsed under running tap water for 10 min, surfacedisinfested for 3 min in 3.5% sodium hypochlorite and then rinsed twice in sterile water for 1 min each. Leaf material was macerated in a droplet of sterile water and streaked onto yeast-extractdextrose-calcium-carbonate (YDC) agar (Schaad and Stall, 1988). Plates were incubated at 25 oC. Following 72 hr incubation, yellow-pigmented colonies typical of Xanthomonas spp. were purified on YDC agar by a series of single colony transfers. Production of brown diffusible pigment on YDC differentiated Xapf from Xap isolates (Basu and Wallen, 1967). Agglutination of antiserum specific to Xap and Xapf, obtained from Adgen Agrifood Diagnostics, Auchincruive, Scotland, was used to identify isolates. Pathogenicity tests on susceptible cultivar Teebus were done to confirm identity of isolates. Pathogenicity tests. Seed from eight tepary lines previously reported to differentiate between Xap and Xapf races (Table 1) (Opio et al., 1996), were obtained from Dr. DP Coyne, University of Nebraska, Lincoln, USA and multiplied from a single seed in a greenhouse to ensure genetically uniform material. The tepary differential set was expanded to include resistant genotypes, XAN 159, GN #1 Nebr. sel 27, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6. Resistance in these lines are all tepary derived. Cultivar Teebus was included as susceptible check. Five seeds of each genotype were planted in 15-cm-diameter plastic pots in sterile soil and maintained in a greenhouse at 18 oC night/28 oC day. Seedlings were thinned to four plants per pot after emergence. One pot per differential was used per isolate, each plant representing a replicate. Pots were randomised prior to inoculation. Experiments were repeated twice to confirm reactions of isolates. One hundred and fourty three isolates from 44 localities in four countries of southern Africa were selected for the study (Table 2a, 2b, 2c). Four isolates received from the International Centre for Agriculture in the Tropics (CIAT) were included as reference cultures. Isolates used for each experiment were regenerated from storage at -72 oC, because loss of pathogenicity was encountered by sub-culturing. Inoculum was prepared by suspending 48 to 72-h-old cultures in sterile distilled water and adjusting it turbidimetrically to contain approximately 108 CFU.ml-1. Fourteen to 20-day-old plants with fully expanded first trifoliate leaves were used for inoculation. Plants were inoculated using the multiple-needle inoculation method (Andrus, 1948). Control plants were inoculated with sterile distilled water. Inoculated plants were kept in a greenhouse at 18 oC night/28 oC day. Plants were rated for infection 14 days after inoculation on a 1 to 9 scale (Aggour et al., 1989). Plants rated 1 to 3, were classified as resistant (incompatible) and ratings of 4 to 9 considered susceptible (compatible). Isolation of bacterial DNA. Eight isolates (two Xap and six Xapf) from southern Africa, one Xapf isolate from CIAT and a non-pathogenic Xanthomonas isolate (Table 3) were used in genetic studies. These isolates were selected based on their geographic origin. Isolates were cultured in 50 ml nutrient broth for 24-48 hr at 25 oC prior to DNA isolation. Bacterial cells were collected by centrifugation at 5 000 rpm for 10 min. Cells were washed three times by resuspending in 5 ml 1 M NaCl and centrifugation at 5 000 rpm for 10 min; followed by two wash steps in 5 ml sterile distilled water. Washed cells were resuspended in 10 ml warm (55 oC) extraction buffer containing 0.2 M Tris.HCl (tris (hydroxymethyl) aminomethane), pH 8.0; 10 mM EDTA (ethylenediaminetetraacetate), pH 8.0; 0.5 M NaCl; 1% (w/v) SDS (sodiumdodecylsulfate) and 10 mg.ml-1 Proteinase K. Resuspended cells were incubated in a water bath at 55 oC for one hr and half a volume 7.5 M ammonium acetate was added. The suspension was mixed by gentle inversion and incubated at room temperature for 10 min. Phase separation was enhanced by adding 100 ml TE buffer (10 mM Tris.HCl, pH 8.0; 1 mM EDTA, pH 8.0). Phases were separated by centrifugation at 14 000 rpm for 15 min. The upper aqueous layer was transferred to a fresh tube containing an equal volume of isopropanol, mixed by gentle inversion and incubated at room temperature for a minimum of 2 hr to overnight. DNA was collected by centrifugation at 14,000 rpm for 15 min. The precipitated DNA was washed twice in 1 ml ice-cold 70% (v/v) ethanol, the pellet air-dried at room temperature, and resuspended in 10 ml TE buffer. The DNA was treated with RNase for two hours at 37 oC and concentration and purity estimated by measuring absorbances at A260 and A280. DNA samples were diluted to a working solution of 200 ng ml-1 (Sambrook et al., 1989). RAPD analysis. Arbitrary 10 bp oligonucleotide primers (Operon Technologies, Table 4) were used for the polymerase chain reaction (PCR) based on the protocol of Williams et al. (1990), with minor modifications. Amplification reactions were performed in a 25 ml reaction volume containing Promega (Promega Corporation, Madison, Wisconsin) reaction buffer (500 mM KCl; 100 mM Tris.HCl, pH 9.0 at 25°C; 1% (v/v) Triton X-100), 2 mM MgCl2, 100 mM of each dNTP (dATP, dCTP, dGTP and dTTP), 5 pmol primer, 0.5 units Taq DNA polymerase (Promega) and 25 ng template DNA. Reactions were performed using a Hybaid Thermal Cycler (Hybaid Limited, UK) programmed for 5 min at 95 °C, 55 cycles of 1 min at 95 oC, 1.5 min at 35 oC, and 2.5 min at 72C, followed by one cycle of 5 min at 72.5 °C and 5 min at 28 °C. The amplification products were analysed by electrophoresis on 1.5% (w/v) agarose gels (Seakem LE) at 80V for 2 hr using UNTAN buffer (0.4 M Trisbase; 0.02 M EDTA, pH 7.4) and detected by staining with 1 mg ml-1 ethidium bromide. Gels were photographed under UV light with polaroid 667 film. All reactions were repeated and only reproducible bands were considered in this study. AFLP analysis. AFLP adapters and primers were designed based on the method of Vos et al. (1995). Adapter and primer sequences are given in Table 5. Primers were synthesised by GibcoBRL (Life Technologies, Glasgow, United Kingdom) and oligonucleotides used for the adapters were PAGE (polyacrylamide gel electrophoresis) purified. Adapters were prepared by adding equimolar amounts of both strands, heating for 10 min to 65 oC in a water bath and leaving it to cool down to room temperature. AFLP adapters and primers were designed based on the methods of Vos et al. (1995). Adapter and primer sequences are given in Table 5. Primers were synthesised by GibcoBRL (Life Technologies, Glasgow, United Kingdom) and oligonucleotides used for the adapters were PAGE (polyacrylamide gel electrophoresis) purified. Adapters were prepared by adding equimolar amounts of both strands, heating for 10 min to 65oC in a water bath and leaving it to cool down to room temperature. AFLP analysis was performed following the protocol described by Vos et al. (1995) and the product manual supplied by Life Technologies Inc. (Glasgow, UK), with minor modifications. Restriction enzymes EcoRI and MseI were used to digest 500 ng of isolate genomic DNA for 4 hr and the reaction mixture, without inactivation of the restriction endonucleases, was subjected to the overnight ligation of adapters at 37 °C, followed by pre-amplification. The ligation mixture was not diluted prior to pre-amplification and the pre-amplification DNA was diluted only 1:5 prior to selective amplification. The selective amplification was conducted using two primers, and the MseI primers always had three selective nucleotides while the EcoRI primers had two, three or four selective nucleotides (Table 5). Gel electrophoresis. Gel electrophoresis for AFLP analysis was performed using the protocol of Vos et al. (1995) but employing a 5% (w/ v)denaturing polyacrylamide gel (19:1 acrylamide: bis-acrylamide; 7 M urea; 1x TBE buffer (89 mM Tris-borate; 2.5 mM EDTA)). Electrophoresis was performed at constant power, 80 W for approximately 2 hr. Silver staining for DNA visualisation. Polyacrylamide gels were silver-stained following the protocol described by the Silver SequenceTM DNA Sequencing System manual supplied by Promega (Madison, WI, USA). The gels were left upright overnight to air dry and photographed by exposing photographic paper (Kodak Polymax II RC) directly under the gel to about 20 sec of dim light. This produced a negative image, exactly the same size of the gel. Statistical analyses. Data obtained from RAPD and AFLP analysis on ten isolates were used for statistical analysis. DNA bands obtained for each isolate were scored based on their presence (1) or absence (0). Only reliable and repeatable bands were considered. Pair wise genetic distances were calculated between isolates Nei and Li (1979). Cluster analysis was done by the unweighed paired group method using arithmetic averages (UPGMA). All calculations were done with the aid of the programme NTSYSpc version 2.02i. RESULTS Identification of isolates. All isolates collected (except Z93) were identified as Xap and Xapf on the basis of their agglutination of specific antiserum and pathogenicity on cv. Teebus (Table 2a, 2b, 2c). Isolate, Z93 did not induce any disease on cv. Teebus and exhibited a weak reaction when tested with the antiserum. The majority of isolates (72%) produced a brown diffusible pigment on YDC agar and were classified as Xapf. Differences in aggressiveness between isolates on the cv. Teebus were detected with mean ratings ranging from moderately to highly susceptible (5-9). The most aggressive isolates included both Xap and Xapf. Pathogenicity tests. All isolates inoculated onto the tepary differential set induced reaction on genotype Nebr. #21. The majority of isolates (99,3%) exhibited an incompatible reaction (rating 1-3) on the remaining genotypes, resembling the infection pattern of race 2 (Opio et al. 1996) (Table 1). One isolate (X539) induced disease (mean ratings 4-9) on all tepary genotypes and did not resemble any infection pattern previously reported (Table 1). A small percentage of isolates induced a slight reaction on genotypes Nebr. #1 (6.3%; rating=1-2.25), Nebr. #5 (1.4%; rating=12.3), Nebr. #8b (9.1%; rating=1-2.0), Nebr. #19 (1.4%; rating=1-1.5), PI 321638 (23.1%; rating=12.8) and L242-45 (4.2%; rating=1-1.5). These reactions were not repeatable in further experiments and reactions were, therefore, considered incompatible with mean ratings not exceeding 3. No symptoms developed on Nebr. #22 except when inoculated with isolate X539. Teebus was susceptible to all the isolates tested except for one non-pathogenic isolate (Z93) that did not induce disease on any of the inoculated lines. Genotypes XAN 159, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6 were generally resistant to all isolates (mean rating=1-3). Six isolates (X563, X573, X121, X295, X561 and X594) induced disease on XAN 159 with a mean rating of 4. GN #1 Nebr. sel 27 were susceptible to all isolates (mean rating=7). RAPD analysis. RAPD analysis produced between two and ten fragments (Fig. 1), but results were not repeatable as a result of sensitivity to variable conditions in laboratory. Best results were obtained with primer OPA-02. RAPD analysis revealed a high frequency of DNA polymorphism among isolates and were able to distinguish between Xap, Xapf and the nonpathogenic isolate. AFLP analysis. The AFLP fingerprinting techniques revealed complex banding patterns that were difficult to interpret due to complex banding patterns. (Fig. 2). DNA fingerprinting techniques revealed a high frequency of DNA polymorphism among isolates with a low presence of shared fragments between isolates (Fig. 2). A total of 756 fragments were amplified using 16 primer pair combinations. Only 2.64% of these fragments were shared between all ten isolates. Primer combinations varied in their ability to detect polymorphisms, ranging from 16 to 86 polymorphisms per primer pair, with an average of 47.3 fragments per primer combination. Fragment sizes varied between 100 and 900 base pairs. Selectivity of AFLP analysis, using two restriction enzymes, was enhanced, by using primers containing two, three or four selective nucleotides. This enhancement of primer selectivity did not reduce the complexity of resulting AFLP banding patterns. Best results were obtained when primers containing three selective nucleotides were used in the AFLP analysis. As with RAPD analysis, the AFLP technique also separated Xap, Xapf and the non-pathogenic isolate into different groups. Fingerprinting techniques, thus, clearly differentiated amongst Xap as well as Xapf isolates. Combined data produced by RAPD and AFLP techniques are shown in Figure 3. The phenogram drawn using pooled data from the RAPD and AFLP analysis (Fig. 3), showed a maximum similarity between any two isolates of 81% (Xapf isolates Les19 and Xapf180). The minimum similarity between any two isolates was 67.5% (Xap isolates X448 and X590). The Xapf cluster of isolates was linked to the Xap cluster of isolates at a similarity of 45.6% and the non-pathogenic isolate Z93 was linked to the Xapf/Xap cluster with a similarity of 30.6%. Isolates within the Xapf cluster exhibited a similarity of 71%. The obtained cophenetic correlation (r=0.994) indicated that the UPGMA cluster analysis was statistically significant. DISCUSSION Results of this study, based on pathogenicity and molecular characterisations, showed that diversity exists within Xap(f) populations, in southern Africa (Table 1, Fig. 1, Fig. 2, Fig. 3). Isolates differed in production of brown pigment as well as aggressiveness on the cv. Teebus. Although it has previously been reported that pigment producing Xapf isolates are more aggressive (Leakey, 1973; Ekpo and Saettler, 1976; Bozzano-Saguier and Rudolph, 1994; Opio et al., 1996), the most aggressive isolates in this study included both Xap and Xapf. Isolates with lower levels of aggressiveness, however, belonged to Xap (rating on cv Teebus=5). Gilbertson et al. (1991) and Tarigan and Rudolph (1996) reported that pigment is not associated with pathogenicity and should be considered of little pathological importance (Schuster and Coyne 1975). Although no differences in disease reaction were observed, RAPD and AFLP analyses demonstrated that Xap and Xapf represent two distinct groups of bacteria. All isolates (except X539 and Z93) inoculated on the tepary differential set had an identical infection pattern, similar to race 2 following the classification of Opio et al. (1996). Although a number of isolates induced only a mild reaction on some of the tepary lines, these reactions were not always repeatable, which is similar to results obtained by Zaiter et al. (1989). The reason for the slight reaction is unknown but may be due to slight variation in greenhouse conditions. The slight reaction was, however, still rated as incompatible (rating <3). The nonpathogenic isolate (Z93) did not induce disease on any of the lines tested. All isolates (except X539 and Z93) inoculated on the tepary differential set had an identical infection pattern, similar to race 2 following the classification of Opio et al. (1996). Although a number of isolates induced only a mild reaction on some of the tepary lines, these reactions were not always repeatable, which is similar to results obtained by Zaiter et al. (1989). The non-pathogenic isolate (Z93) did not induce disease on any of the lines tested. Except for isolate X539, which exhibited a significantly different infection pattern, no races other than race 2, previously described by Opio et al. (1996), could be distinguished. The distinct pattern of differential reaction recorded for this isolate, may represent another, as yet unrecorded, race of Xap. The possibility exists that isolates identical to X539 exist, but may not have been sampled in this study. Continuous monitoring of CBB isolates in future is necessary in order to detect presence of isolates exhibiting differential reactions. Although isolate X539 was pathogenic on the eight tepary lines tested, no disease developed on resistant genotypes used to supplement the differential set, except for GN #1 Nebr. sel. 27. Using these resistant genotypes in a resistance breeding programme would, therefore, not be influenced by the occurrence of this isolate. Except for isolate X539, which exhibited a significantly different infection pattern, no races other than race 2, previously described by Opio et al. (1996), could be distinguished. The distinct pattern of differential reaction recorded for this isolate, may represent another, as yet unrecorded, race of Xap. The possibility exists that isolates identical to X539 exist, but may not have been sampled in this study. Continuous monitoring of CBB isolates in future is necessary in order to detect presence of isolates exhibiting differential reactions. Although isolate X539 was pathogenic on the eight tepary lines tested, no disease developed on resistant genotypes used to supplement the differential set, except for GN #1 Nebr. sel. 27. Using these resistant genotypes in a resistance breeding programme would, therefore, not be influenced by the occurrence of this isolate. Genotypes XAN 159, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6 that were used to supplement the tepary differential set (Table 1), were generally resistant to all isolates tested. Resistance in all these lines is tepary-derived. XAN 159 was slightly susceptible to a small number of isolates. Resistance instabilities such as these have been reported previously in XAN 159 and its progeny (Beebe and Pastor-Corrales 1991), however, it is still widely used in resistance breeding programmes (Beebe and Pastor-Corrales, 1991; Fourie and Herselman, 2002; Park et al., 1998; Mutlu et al., 1999; Singh and Muòoz, 1999). Genotypes XAN 159, Wilk 2, Wilk 6, Vax 4, Vax 5 and Vax 6 that were used to supplement the tepary differential set, were generally resistant to all isolates tested. Resistance in all these lines is tepary-derived. XAN 159 was slightly susceptible to a small number of isolates. Resistance instabilities such as these have been reported previously in XAN 159 and its progeny (Beebe & Pastor-Corrales 1991), however, it is still widely used in resistance breeding programmes (Beebe & Pastor-Corrales 1991, Fourie & Herselman 2002, Park et al. 1998, Mutlu et al. 1999, Singh & Mu oz 1999). The reportedly resistant line GN #1 Nebr. sel 27 (Coyne and Schuster, 1983)was susceptible to all the isolates used in this study. This line was originally derived from inter-specific crosses between P. vulgaris and P. acutifolius and has been used in many breeding programmes as a source of resistance (Coyne and Schuster, 1974; Mohan and Mohan, 1983). Recent molecular studies have, however, indicated that resistance in GN #1 Nebr. sel 27 is derived from P. vulgaris and not P. acutifolius, as previously described (Miklas et al., 2003). Although susceptible in South Africa, GN #1 Nebr. sel 27 and lines derived from it, have tested resistant in some parts of the USA (Coyne and Schuster, 1974). The reportedly resistant line GN #1 Nebr. sel 27 was susceptible to all the isolates used in this study. This line was originally derived from inter-specific crosses between P. vulgaris and P. acutifolius and has been used in many breeding programmes as a source of resistance (Coyne & Schuster 1974, Mohan & Mohan 1983). Recent molecular studies have, however, indicated that resistance in GN #1 Nebr. sel 27 is derived from P. vulgaris and not P. acutifolius, as previously described (Miklas et al. 2002). Although susceptible in South Africa, GN #1 Nebr. sel 27 and lines derived from it, have tested resistant in some parts of the USA (Coyne & Schuster 1974) and Spain (C. Assensio, MBGCSIC: personal communication). Inconsistency in these results could have resulted from the limited distribution of Xapf in some areas of the USA and Spain (R. Gilbertson, University of California-Davis: personal communication). Results of DNA fingerprinting techniques indicated that genetic diversity exists among isolates of the common blight pathogen (Fig. 1- Fig. 2). Differences between Xap and Xapf isolates show that these represent two distinct groups of bacteria. Similar distinction between these two groups was also reported by Gilbertson et al. (1991), using RFLP’s. Non-pathogenic Xanthomonas commonly associated with beans could be distinguished from Xap and Xapf using both RAPD and AFLP techniques. These results are similar to those of Gilbertson et al. (1990) who distinguished between non-pathogenic and pathogenic isolates using RFLP’s. Results of DNA fingerprinting techniques indicated that genetic diversity exists among isolates of the common blight pathogen. Differences between Xap and Xapf isolates show that these represent two distinct groups of bacteria. Similar distinction between these two groups was also reported by Gilbertson et al. (1991), using RFLP’s. Nonpathogenic Xanthomonas commonly associated with beans could be distinguished from Xap and Xapf using both RAPD and AFLP techniques. These results are similar to those of Gilbertson et al. (1990) who distinguished between nonpathogenic and pathogenic isolates using RFLP’s. Although isolate X539 gave a significantly different infection pattern when inoculated onto the tepary lines, no significant difference between this isolate and the others Xapf isolates could be detected using different molecular techniques. It has been reported that strains of Xap and Xapf from similar geographic locations had similar, but not identical RFLP patterns (Gilbertson et al., 1991; CIAT, 1992). This could not be confirmed in the present, study and is possibly due to the small number of isolates tested. Results obtained in this study indicate that both pathogenic and genetic variation exist in the CBB pathogen population in southern Africa. However, identical reactions with the majority of isolates on the tepary lines, showed that different CBB races do not occur. Information gained from this study made it possible to select the most appropriate isolates to use in a resistance breeding programme. Results obtained in this study indicate that both pathogenic and genetic variation exist in the CBB pathogen population in southern Africa. However, identical reactions with the majority of isolates on the tepary lines, showed that different CBB races do not occur. Information gained from this study made it possible to select the most appropriate isolates to use in a resistance breeding programme. ACKNOWLEDGEMENT Partial support for this work provided by the Pan-Africa Bean Research Alliance (PABRA) is highly appreciated. REFERENCES
Copyright © 2011, African Crop Science Society The following images related to this document are available:Photo images[cs11037f2.jpg] [cs11037t2a.jpg] [cs11037t3.jpg] [cs11037t1.jpg] [cs11037t5.jpg] [cs11037t4.jpg] [cs11037f1.jpg] [cs11037t2b.jpg] [cs11037f3.jpg] [cs11037t2c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}