 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 20, No. 1, March, 2012, pp. 17-29 Induction of oxidative stress and antioxidative mechanisms in hyacinth bean under zinc stress R. Myrene D’Souza And V.R. Devaraj1 CSIR-SRF, Department of Biochemistry, Central College Campus, Bangalore University,
Bangalore, 560 001,Karnataka, India
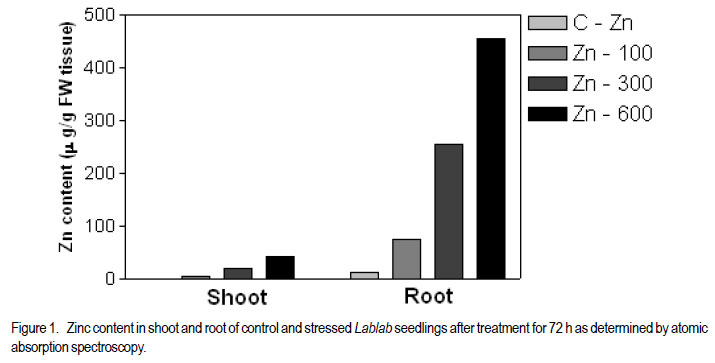
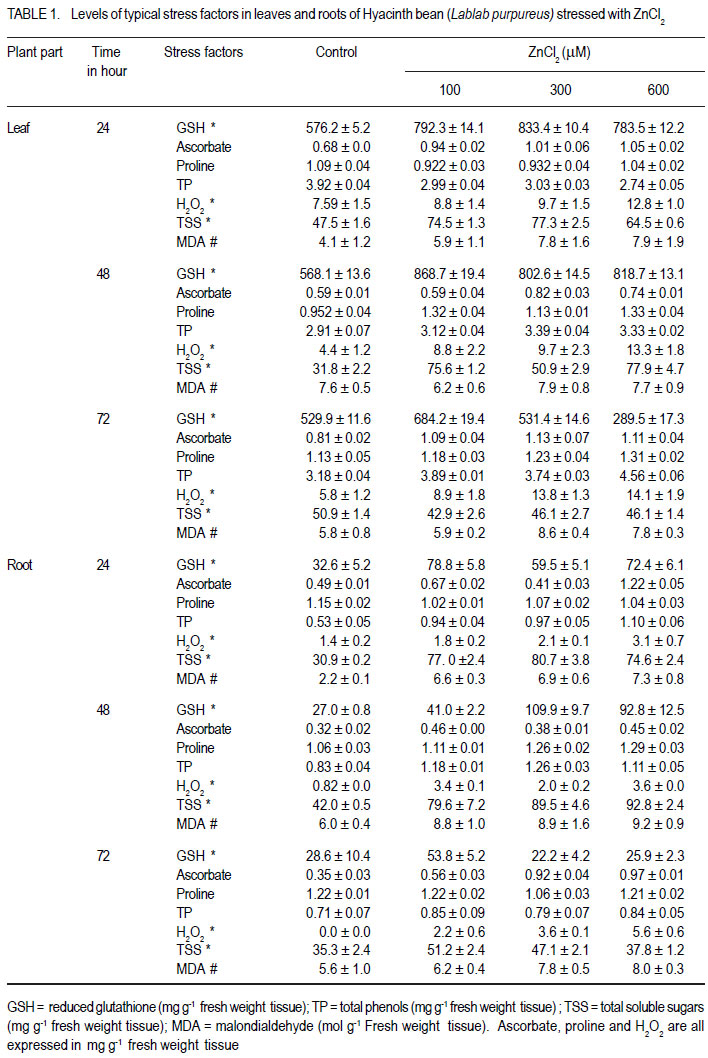
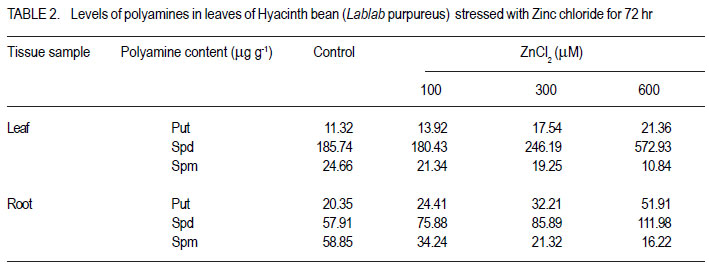
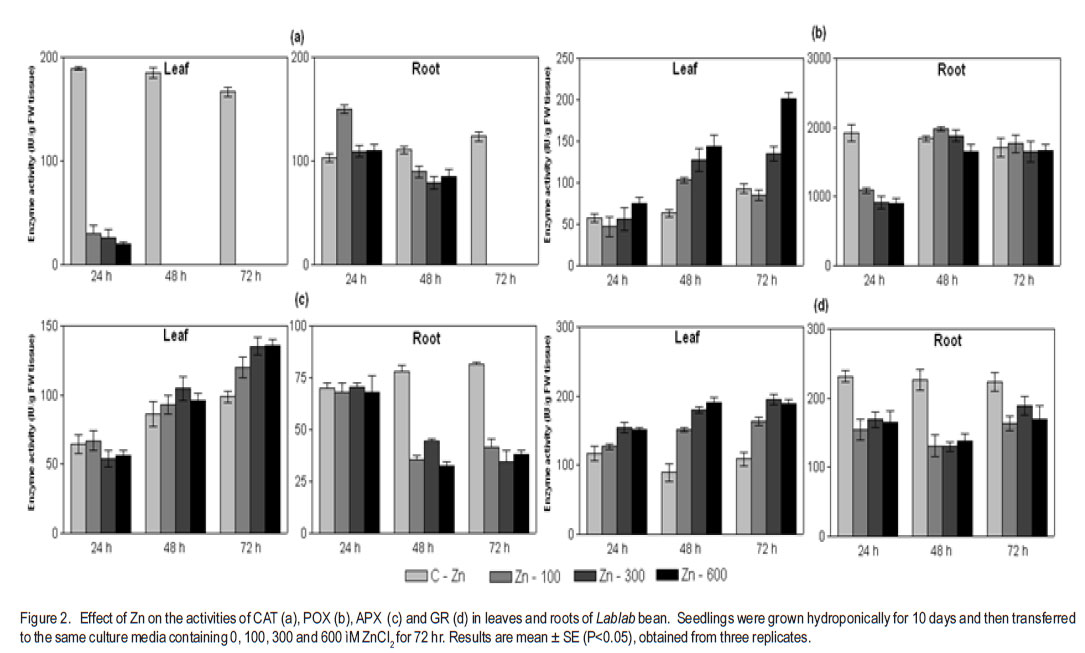
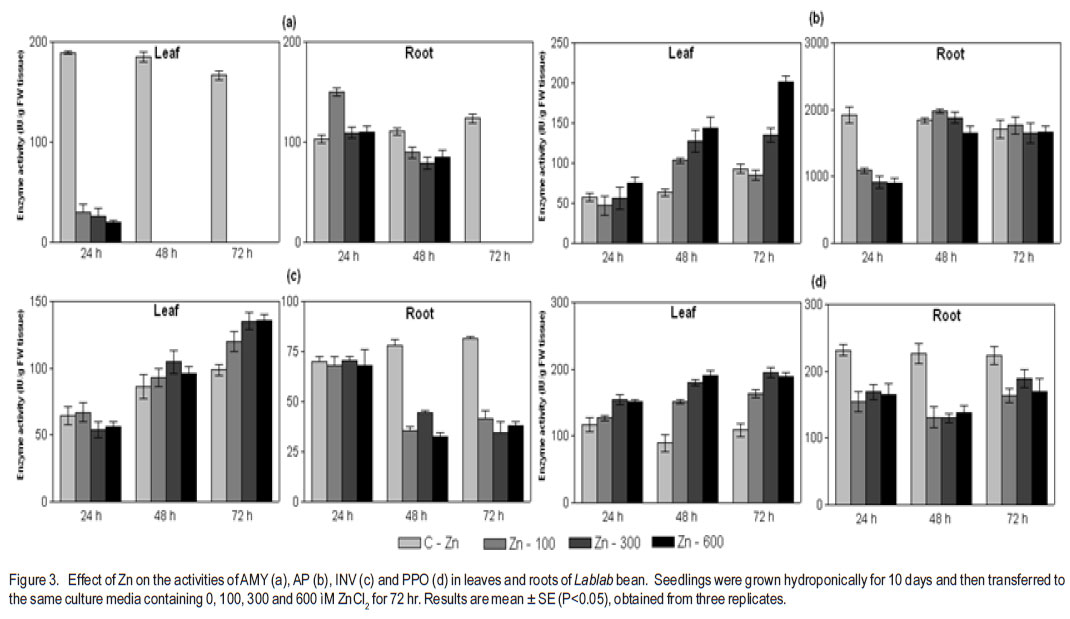
1Department of Biochemistry Central College Campus, Bangalore University, Bangalore, 560001, India Recieved 25 July, 2011; accepted 31 January, 2012 Code Number: cs12002 ABSTRACT Zinc toxicity to plants occurs in soils contaminated by mining and smelting activities, in agricultural soils treated with sewage sludge, and in urban and peri-urban soils enriched by anthropogenic inputs of Zn. Zinc-induced stress in Hyacinth bean (Lablab purpureus) was investigated by growing seedlings in hydroponics, supplemented with Zinc (0-600 µM), under controlled growth conditions. Changes in growth parameters, enzyme activities and other stress response factors, as a function of Zn uptake are directly or indirectly related to the cellular free radical scavenging systems. These parameters were studied in 10-day old Lablab seedlings over 72 h of exposure. Relative to leaves, roots accumulated ~12-fold higher amounts of Zn. Oxidative stress markers, H2O2, malondialdehyde, ascorbate, proline, soluble sugars, putrescine and spermidine were elevated in both tissues, whereas, glutathione and spermine levels showed a steady decline. Total phenol increased in roots, but decreased in leaves. Zn-stress enhanced antioxidant enzymes, guaiacol peroxidase, glutathione reductase, and polyphenol oxidase in concentration and time dependent manner in leaves, but showed an inverse relationship in roots. Catalase decreased in both leaves and roots. Metabolic enzymes β-amylase and acid phosphatase increased in both tissues. Invertase increased in leaves but declined drastically in roots. The results suggest that primary antioxidative response originates in leaves of Lablab, while roots are involved in direct uptake of heavy metals and are not adept in extrusion. Also, lower concentrations of Zn (up to 100 µM) stimulate growth of Lablab, but higher concentrations proved detrimental to the plant. Key Words: Lablab purpureus, Malondialdehyde, Polyamines RÉSUMÉ Les effets toxiques du Zinc aux cultures se produit dans des sols contaminés par des activités d’exploitation et extraction, dans des sols avec dépots d’égouts, et en sols urbains et peri-urbains enrichis en depots anthropogéniques riches en Zn. Une recherche sur le stress dû au zinc était effectuée sur des plantules du Hyacinth (Lablab purpureus) en conditions hydroponiques avec supplément du Zinc (0-600 mM) en conditions de croissance controllée. Les changements des paramètres de croissance, des activités enzymatiques et autres facteurs de réaction au stress, par suite de l’importation du Zinc sont directement ou indirectement en rapport avec des systèmes radicaux cellulaires de fouille libre. Ces paramètres étaient étudiés sur des plantules de lablab de 10 jours exposées pendant 72 heures. En comparaison avec les feuilles, les raciness ont accumulé 12 fois plus le contenu en Zn. Les marqueurs oxydatifs de stress, H2O2, la malondialdehyde, l’ascorbate, la proline, les sucres solubles, la putrescine et la spermidine étaient élevés dans les tissus, alors que les niveaux de la glutathione et la spermine ont montré une baisse continue. Le phenol total a augmenté dans les raciness, mais décru dans les feuilles. Le stress dû au Zinc a accru la concentration d’enzymes antioxydants, le peroxidase guaiacol, le reductase glutathione, et l’oxidase polyphenol dans les feuilles, mais a montré une relation inverse dans les racines. Par ailleurs, le catalase a décru dans les feuilles et racines. Les enzymes métaboliques b-amylase et le phosphatase acide ont augmenté dans les tissus, tandis que l’invertase a augmenté dans les feuilles et décru drastiquement dans les racines. Les résultats ont suggéré que la réaction antioxydante primaire provient des feuilles de Lablab, pendant que les racines sont impliquées dans des exportations directes des métaux lourds et ne sont pas sujettes à l’ extrusion. Aussi, des plus basses concentrations du Zn (jusqu’100mM) ont stimulé la croissance du lablab pendant que des concentrations les plus élevées ont induit des effets nuisibles à la proline de la plante. Mots Clés: Lablab purpureus, Malondialdehyde, Polyamines INTRODUCTION Elevated levels of both essential and nonessential heavy metals in the soil result in decreased growth and productivity of higher plants. Recent literature on heavy metal stress, indicates a dynamic action similar to other stress factors. The toxicity of heavy metals proceeds in two stages. The first inductive stage (lasting 1-2 days) involves the heightened activity of signaling processes, enzyme activity and changes in structures of cell wall and cell membrane. The second stage (lasting over 2 days) entails prolonged and permanent effects that include biochemical changes allowing diminution of stress and degradation processes leading to senescence (Maksymiec et al., 2008). Zinc is a natural constituent of the earth’s biogeochemical cycles, and is required by plants in trace amounts. As a result of human activities such as metal mining, the use of agrochemicals, and the agricultural use of sewage sludge, Zn can be present in soils in excessive amounts, resulting in the need for genotypes that are more tolerant to high levels of Zn. When present in excess in plant tissues, it is extremely toxic to plant cells. High concentration of Zn can affect ionic homeostatic systems by interfering with the uptake, transport and regulation of essential ions. It results in disruption of metabolic processes such as transpiration (Rout and Das, 2003), photosynthesis (Bonnet et al., 2000) and metabolic enzyme activity (Van Assche et al., 1988). Very high levels of Zn may also competitively inhibit the function of metalrequiring proteins and form complexes with DNA and RNA decreasing their stability (Broadley et al., 2007). Elevated levels of heavy metals in plant tissue are known to generate reactive oxygen species (ROS). Redox-active metals (e.g. Cu and Fe) participate in the Haber-Weiss reaction and produce hydroxyl radicals; while metals without redox capacity (e.g. Cd, Hg and Zn) cause metabolic imbalance, disrupt the antioxidative potential of the cell, and eventually enhance the ROS load (Briat, 2002). Due to transition property, Zn ions are able to induce oxidative stress by triggering generation of free radicals and ROS such as, H2O2, superoxide and hydroxyl radical, formed by the leakage of electrons onto molecular oxygen from the electron transport chain of mitochondria and chloroplasts during normal metabolism. To eliminate the harmful effects of ROS, plant cells are equipped with a well-developed antioxidant defense system comprising enzymes; namely, catalase (CAT), guaiacol-specific peroxidase (POX) and glutathione reductase (GR), ascorbate peroxidase (APX) as well as nonenzymatic components such as glutathione (GSH), ascorbate (ASC), proline (PRO), phenols and polyamines. Legume crops are reported to be tolerant to several heavy metals. There has been considerable interest in finding legume species that are able to colonise metal-enriched soils for use in land reclamation (Day and Ludeke, 1981). Lablab purpureus (Hyacinth bean) is extensively cultivated in India, Africa and Indonesia. This paper attempts to evaluate the ability of Hyacinth bean to tolerate toxic levels of Zn in terms of biochemical parameters such as levels of antioxidants, antioxidant enzymes and other markers of abiotic stress. MATERIALS AND METHODS Plant material and zinc treatment. One hundred grammes of seeds of Lablab bean obtained from the National Seed Project, University of Agricultural Science, Bengaluru, India, were surface sterilised with 0.1% (w/v) mercuric chloride for 1 min, rinsed immediately with about 3 litres of distilled water and imbibed overnight in distilled water. The overnight-soaked seeds were sown in trays containing vermiculite and acid-washed sand (1:1 w/w), and irrigated daily with 10 ml distilled water per pot. The plants were exposed to natural photoperiod of 12 hr, with a temperature of 28 ± 4 °C. Ten-day old seedlings were subjected to Zn stress with 100, 300 and 600 µM ZnCl2 in halfstrength Hoagland medium (Hoagland et al., 1950). Ten seedlings were used per concentration regime. Leaf and root samples were collected 24, 48 and 72 hr after application of stress; and frozen until further analysis. The experiment was conducted in a completely randomised design with 3 evaluation points (24, 48 and 72 hr after stress). Each set up was replicated three times. Metal uptake. Total leaf and root accumulation of Zn in Lablab bean was determined after 72 hr of treatment. Leaves and roots were harvested, washed in deionised water for 2 mins, air-dried and oven-dried at 80 ºC for 48 hr. The tissue was ground into a fine powder using a chilled porcelain pestle and motar. One gramme of this powder was dissolved in binary mixture containing 3 parts of 1M HNO3 and 1 part of 1M HCl, and the metal concentration was analysed by flame photometery (Systronics India Limited, Model No. 128). Zinc chloride at different concentrations (0.1 to 2.0 µg ml-1) was used as the standards. Determination of stress response factors. Hydrogen peroxide contents in the control and stressed seedlings were determined according to Velikova et al. (2000). Leaf and root tissues (500 mg) were homogenised separately in an ice bath with 5 ml of 0.1% (w/v) trichloroacetic acid. The homogenate was centrifuged at 10,000 rpm for 15 min, and 0.5 ml of the supernatant was added to 0.5 ml of 10 mM potassium phosphate buffer (pH 7.0) and 1 ml of 1 M potassium iodide . The absorbance of the supernatant was measured at 390 nm. Glutathione (GSH) was estimated according to Beutler (1963). The tissue was homogenised with 3% metaphosphoric acid. DTNB [5,5'dithiobis(2-nitrobenzoic acid)] was added to supernatants cleared by centrifugation. The formation of 5-thio-2-nitrobenzoic acid, which is proportional to total glutathione concentration, was monitored at 412 nm at 25 °C against reagent controls (Beutler, 1963). Ascorbic acid estimation was carried out according to the procedure of Sadasivam and Manickam (1997). The tissue was homogenised in 4% oxalic acid and centrifuged at 10,000 g for 10 min. The assay mixture consisted of 0.1 ml of brominated sample extract made up to 3.0 ml with distilled water, 1.0 ml of 2% 2,4dinitrophenylhydrazine reagent and 1-2 drops of thiourea. After incubation at 37 ºC for 3 hr, the orange-red osazone crystals formed were dissolved by the addition of 7.0 ml of 80% sulphuric acid and the absorbance was read at 540 nm. Proline content was estimated using ninhydrin reagent according to Bates et al. (1973). The amount of total soluble sugars was estimated colorimetrically at 540 nm, using anthrone reagent according to Roe (1955). Total phenols were estimated by the method of Slinkard and Singleton (1977), using catechol as an authentic standard. The amount of total soluble sugars was estimated colorimetrically at 540 nm using anthrone reagent as mentioned in Roe et al. (1955). The extent of lipid peroxidation was determined according to Heath and Packer (1968), with suitable modification. In this case, 0.5 g of fresh tissue was ground in 5.0 ml of 0.1% thiobarbituric acid (TCA) containing 0.5% butylated hydroxytoluene containing 1.0% PVP. The homogenate was centrifuged at 12,000 g for 30 min. Four millilitres of the supernatant was mixed with 4.0 ml of the substrate (0.5% thiobarbituric acid and 20% TCA). The mixture was boiled for 30 min, chilled on ice, and centrifuged at 12,000xg for 10 min. The absorbance of supernatant at 532 nm was measured and the nonspecificabsorbance at 600 nm was subtracted. The MDA content was calculatedfrom the extinction coefficient of 155 mM-1 cm-1. Polyamines were detected by HPLC according to the procedure mentioned in Redmond and Tseng (1979) with slight modifications. One ml of 2 M NaOH was mixed with 500 µl of HClO4 extract. After addition of 10 µl benzoyl chloride, vortexing for 20 s, and incubation for 20 min at 37 °C; 2 ml saturated NaCl was added. Benzoyl-polyamines were extracted in 2 ml diethyl ether by adding the solvent to the mixture followed by brief vortexing. After centrifugation at 1500 × g for 5 min at 4 °C, 1 ml of the ether phase was collected, evaporated to dryness under a stream of warm air, and redissolved in 100 µl methanol. Standards were treated in a similar way. The benzoylated samples were filtered through 0.22 µm membrane filter, then eluted at room temperature on a reverse-phase C18 column employing isocratic elution with methanol:water (64:36 v/v) system at a flow rate of 1 ml min-1. Extraction of enzymes. The frozen samples were homogenised with pre-chilled 50 mM sodium phosphate buffer (pH 7.0), containing 5 mM βmercaptoethanol and 1 mM EDTA using pestle and mortar. The homogenate was centrifuged at 12,000 × g for 15 min at 4 °C. The supernatant was used as a source of enzymes. Soluble protein content was determined according to Lowry et al. (1951) using BSA as the standard. Assay of antioxidant enzymes Guaiacol Peroxidase (POX) (E.C. 1.11.1.7). Guaiacol peroxidase activity was measured in a reaction mixture of 3.0 ml consisting of 50 mM phosphate buffer (pH 7.0) containing 20 mM guaiacol, 10 mM H2O2 and 100 µl enzyme extract (Chance and Maehly, 1955). The formation of tetraguaiacol was followed by an increase in A470 nm (ε = 26.6 mM-1 cm-1). One unit of peroxidase is defined as the amount of enzyme needed to convert 1 µmol of H2O2min-1 at 25 °C. Catalase (CAT) (E.C. 1.11.1.6). Catalase activity was assayed by following the decline in absorbance of H2O2 at 240 nm (ε = 39.4 M-1 cm-1) according to the method of Aebi (1984). The reaction mixture consisted of 50 µl of enzyme extract in 50 mM sodium phosphate buffer (pH 7.0). The reaction was started by addition of H2O2 to a final concentration of 10 mM, and its consumption was measured for 2 min. One unit of activity is defined as the amount of enzyme that catalysesthe oxidation of 1 µmol of H2O2 per min under the assay conditions. Glutathione Reducatase (GR) (E.C. 1.6.4.2). GR activity was determined by monitoring the oxidation of NADPH at 340 nm (ε = 6220 M-1 cm-1) according to the method of Carlberg and Mannervik (1985). The reaction mixture contained 50 mM Tris-HCl buffer (pH 7.5), 3 mM MgCl2, 0.5 mM GSSG, 0.2 mM NADPH and 250 ìl of enzyme extract in a total volume of 1.5 ml. One unit of activity is defined as the amount of enzyme that catalysesthe oxidation of 1 µmol of NADPH per min under the assay conditions. Ascorbate peroxidase (APX) (E.C. 1.11.1.11). The activity of APX was determined spectrophotometrically as described by Allen (1968). The assay mixture contained 50 mM HEPES buffer (pH 7.0), 1 mM EDTA, 1 mM H2O2 , 0.5 mM sodium ascorbate and 50 µl of enzyme extract in a total volume of 2.0 ml. The reaction was initiated by addition of H2O2. The oxidation of ascorbate was followed by a decrease in the A290 (ε = 2.8 mM-1 cm-1). One unit of ascorbate peroxidase is defined as the amount of enzyme necessary to oxidize 1 µmol of ascorbateper min at 25 °C. Polyphenol Oxidase (PPO) (E.C. 1.14.18.1). PPO was assayed spectrophotometrically at 400 nm using tertiary butyl catechol as substrate according to Kanade et al. (2006). The assay mixture consisted of 0.9 ml sodium acetate buffer (pH 5.0), 0.1 ml t-butyl catechol and 0.1 ml enzyme extract. The quinone formed was measured at 400 nm (ε = 1150 M-1cm-1). One unit of enzyme activity is defined as the amount of enzyme that produces 1 µmol of t-butylquinone per minute under the assay conditions. Assay of hydrolytic enzymes β-Amylase (AMY) (E.C. 3.2.1.1). Activity of βamylase was measured using the DNS method (Bernfield, 1955). The reaction mixture consisted 0.5 ml of 2% starch solution in 50 mM phosphate buffer (pH 7.0) and 0.5 ml of enzyme extract. Acid Phosphatase (AP) (E.C. 3.1.3.2). AP activity against p-nitrophenol phosphate was determined by monitoring the release of p-nitrophenol at 410 nm according to Hoerling and Svensmark (1976). Each unit of activity is defined as the number of µmoles of p-nitro phenol released per minute. Invertase (INV) (E.C. 3.2.1.26). Invertase activity was determined by the method of Sridhar and Ou (1972). A total of 4.0 ml reaction mixture containing 0.025 M sodium acetate buffer (pH 5.0), 0.625% sucrose and appropriate volume of enzyme extract was incubated at 37 °C for 24 hr. The reaction was arrested by adding equal volume of DNS reagent. Statistical analysis. The experiment was performed using a completely randomised design in three replicates. Comparisons of means were performed using PrismGraph Version 3.02. Data were subjected to a one-way analysis of variance, and the mean differences were compared by least significant difference (LSD) at 5% level of significance. RESULTS AND DISCUSSION Zinc accumulated in both leaves and roots of Lablab seedlings under Zn stress depended on the concentration of Zn in the growth media (Fig. 1). Furthermore, the Zn content was relatively higher in roots than in leaves. At higher Zn stress (600 µM for 72 hr), roots showed 10-fold higher content of Zn than leaves. Similar distribution was observed earlier in Brassica juncea, where the deposition occurred in the form of simple Zn salts or as complexes with proteins and carbohydrates (Sridhar et al., 2005). The shoot to root concentration ratio suggested that Lablab is not an accumulator of Zn and it lacks an efficient translocation system for Zn transport from root to shoot. Both redox active and non-redox active heavy metals have been reported to induce overproduction of ROS (Gajewska and Sklodowska, 2007). The accumulation of H2O2 could also reflect the oxidative stress and changes of antioxidants in plants. Our results have indicated that Zn is able to trigger excessive production of H2O2 in foliar and root tissues (Table 1). H2O2 is involved in a number of regulatory processes such as cell expansion, development, stomatal closure and programmed cell death (Cheng and Song, 2006). Elevated levels of H2O2 in leaves of Lablab are in conformity with the earlier demonstration of its preferential accumulation in leaves (Chao et al., 2008). As excess of H2O2 causes peroxidation of membrane lipids, it can further lead to oxidative damage via destabilisation of membrane compartments. A common product of lipid peroxidation, MDA has been found to be associated with oxidative stress, and is a sensitive diagnostic index of oxidative injury to plasma membrane. Exposure of Lablab to 300 µM of Zn, even for 24 hr induced a significant increase in MDA in both leaf and root (Table 1). The dose-response experiment demonstrated that enhancement of MDA at 48 hr of exposure to 300 µM Zn was 3.1 and 1.5-folds, respectively, in roots and leaves. A similar response was reported in rice (Verma and Dubey, 2003) and tomato (Chamseddine et al., 2009). Polyamines are known to have a profound effect on plant growth and development and are also involved in protection against environmental stresses (Gill and Tuteja, 2010). They associate with phospholipids of the membrane bilayer and prevent their deterioration under stress conditions (Zhao and Yang, 2008). Among various polyamines, Spd is more closely associated with stress tolerance in plants than Put and Spm (Bouchereau et al., 1999; He et al., 2002). The most abundant of the three polyamines, Spd demonstrated a 2-fold and 1-fold increase in leaves and roots, respectively, after 72 hr at maximum Zn treatment (Table 2). Similar results were reported in wheat (Triticum aestivum L.) stressed with NaCl (El-Shintinawy, 2000). A steady increase in Put and rapid enhancement in Spd (Table 2) suggested that Zn stress caused over expression of Spd synthase. Spd was not converted to Spm, which indicates no influence of Zn stress on Spm synthase. These results are in consonance with NaCl-stressed wheat (Simon-Sarkadi et al., 2002). Thus, individual polyamines have different roles during stress response and Spd could be implicated in enhancing heavy metal tolerance, possibly by exerting an antioxidant activity and/or by metal chelation. Heavy metal-induced ROS accumulation is counteracted by intrinsic antioxidant system of plants comprising a variety of enzymatic scavengers and antioxidants. CAT is a hemecontaining enzyme that catalyses the dismutation of H2O2 into oxygen and water. CAT activity decreased significantly in leaves and roots stressed with Zn (Fig. 2a). These observations are constant with those of Hg-treated cucumber seedlings (Cargnelutti et al., 2006). CAT has low substrate affinities compared to POX and requires simultaneous access of two H2O2 molecules at the active site (Willekens et al., 1997). An alternative mode of H2O2 destruction is via peroxidases that are found throughout the cell and have a much higher affinity for H2O2 than CAT (Noctor and Foyer, 1998). The activity of POX increased at all concentrations after 48 hr of stress in leaves, but was not affected in roots (Fig. 2b). The enhanced POX activity in Lablab thus indicates an efficient detoxification of H2O2. Further, the counterbalancing of CAT and POX suggestes a predominant role of POX in antioxidative mechanism of Lablab. Two major enzymes, APX and GR are involved in the ASC/GSH cycle that operates in chloroplasts, cytoplasm, mitochondria as well as peroxisomes (del Río et al., 2006). In chloroplasts, APX reduces H2O2 using ASC as the electron donor. A study in Cd-treated plants demonstrated that leaf peroxisomes respond by improving the capacity of antioxidative enzymes involved in the ASC/GSH cycle (Palma et al., 2002). Our results indicated that APX activities in leaves increase in the presence of Zn (Fig. 2c), and the early enhancement of APX activities suggests that the Zn-triggered antioxidative capacity mainly employs APX for the removal of H2O2. Efficient working of APX requires rapid regeneration of ASC from dehyrdoascorbate, which in turn, is dependent upon the availability of GSH. To maintain homeostasis, GSH must be generated from oxidised glutathione by GR at the expense of NADPH. In the present investigation, the GR activity was enhanced only in leaves and declined with time in roots (Fig. 2d) suggesting operation of ASC/GSH cycle in leaves. These two important components of ASC/GSH cycle, GSH and ASC were elevated in both leaves and roots of Lablab bean under Zn stress. The first 24 hr of exposure showed only a slight increase of both antioxidants in roots. By contrast, higher accumulation was noted in leaves. The increase in GSH levels correlated well with increased GR (Table 1). However, a significant decline in GSH levels was seen at 72 hr of stress in spite of an increase in activity of GR, indicating the possibility of substantial amount of GSH being employed in metal chelation by the formation of phytochelatins, a polymer of GSH. One of the strategies in abiotic stress tolerance in plants is the over-production and accumulation of osmolytes such as soluble sugars and PRO (Table 1). Lablab exposed to Zn stress exhibited marginal elevation in PRO levels as a function of time (Table 1). It has been reported that metal stress results in water deficit, which in turn, causes PRO accumulation (Sharma et al., 1998), thus contributing to osmotic regulation. The elevated levels of PRO in Lablab suggested induction of osmotic stress under zinc stress. PRO accumulation helps the cell in chelates metal ions, maintaining structural integrity of cytoplasmic proteins, serves as a source of carbon and nitrogen, maintains cytosolic pH, NAD(P)+/NAD(P)H ratio, prevents denaturation of enzymes (Siripornadulsil et al., 2002). The synthesis of PRO bears a direct correlation to carbohydrate levels. The total soluble sugars increased steadily in both leaves and roots up to 48 hr of Zn stress and decreased thereafter (Table 1). The increase in concentration of soluble sugars may be due to starch degradation, inhibition of starch synthesis and inhibition of INV. An elevated level of total soluble sugars is important for energy production, stabilisation of cellular membranes, maintenance of turgor, vitrification of the cytoplasm and signaling, which may account for tolerance to heavy metal stress. Thus, TSS and PRO together seem to protect Lablab against metal induced oxidative and osmotic stress. PPO activity increases during the progression of stress and is a possible tolerance mechanism as demonstrated in peach seedlings subjected to freezing stress (Szalay et al., 2005). Elevated PPO levels in Lablab indicated its possible role during early 48 hr of exposure in leaf tissue. Similar results were demonstrated in Nymphoides peltata under Cd stress (Lavid et al., 2001). Total phenols too behave as powerful antioxidants in plant tissues under abiotic stress, protecting the cell from potential damage and increasing the stability of the cell membrane (Sgherri et al., 2004). The phenol content of the Zn stressed leaves exhibited a time dependent increase (Table 1) suggesting that signals for this secondary metabolitemediated response is elicited under extreme metal stress in Lablab bean. Apart from antioxidant enzymes, which have been indisputably implicated in stress response, there are also reports of induction of metabolite enzymes such as; β- AMY, AP and INV under abiotic stress (Yang et al., 2007). AMY (Fig. 3a) and AP (Fig. 3b) activities increased in both leaves and roots of stressed seedlings. Increased activity of AMY leads to accumulation of total soluble sugars in the stressed plant. AP is known to act under water stress by maintaining a certain level of inorganic phosphate, which can be cotransported with H+ along a gradient of proton motive force (Olmos and Hellin, 1997). The hydrolysis of phosphate esters is an important process in energy metabolism, metabolic regulation and a wide variety of cellular signal transduction pathways of plant cells (Vincent et al., 1992). This indicates a common signaling pathway leading to activation of metabolic and antioxidative enzymes. CONCLUSION Zinc at lower levels acts as a promoter of growth as it is an essential element, but induces oxidative stress with increasing concentration (>100 mM) and prolonged exposure. The shoot to root metal concentration ratio (<1) suggested that Lablab is not an accumulator of Zn. Soluble Zn concentration in bulk soil in most contaminated soils being ~ 4 mM, sustenance of Lablab under Zn stress up to 300 mM in soluble form suggestes that it can survive under Zn stress. The levels of stress-markers; antioxidants, enzymes, phenols and polyamines indicate that the antioxidant response elicited by Zn stress in Lablab overlaps with the response elicited by dehydration or salinity stress, and this response could offer protection to the plant up to 100 mM Zn. ACKNOWLEDGEMENT We acknowledge the Council for Scientific and Industrial Research (CSIR), New Delhi, India for JRF (Award No. 09/039(0080)/2007-EMR-I). REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12002t1.jpg] [cs12002f2.jpg] [cs12002f1.jpg] [cs12002t2.jpg] [cs12002f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}