 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
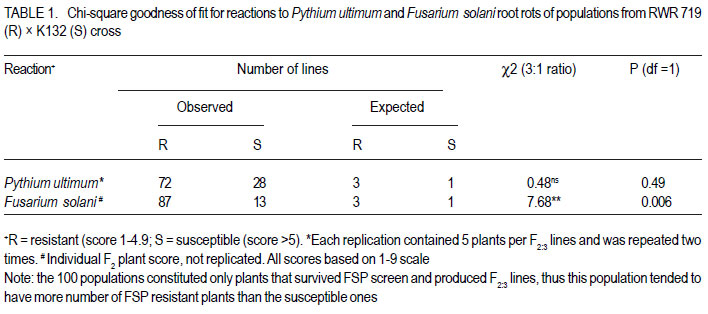
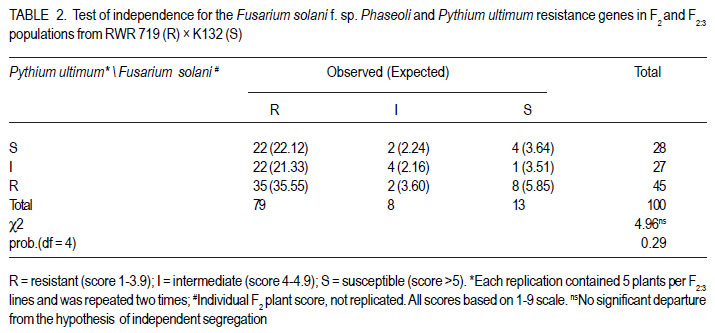
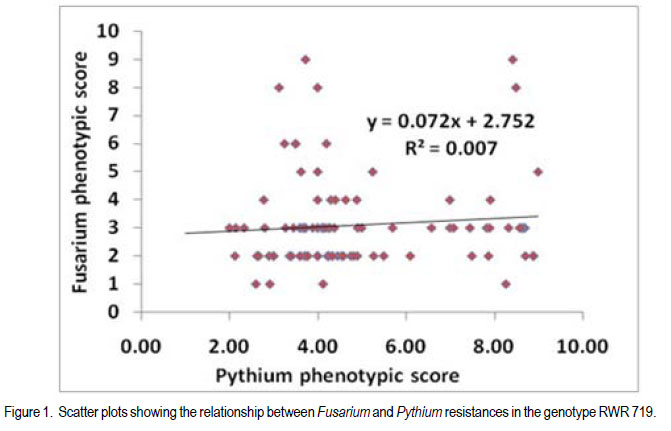
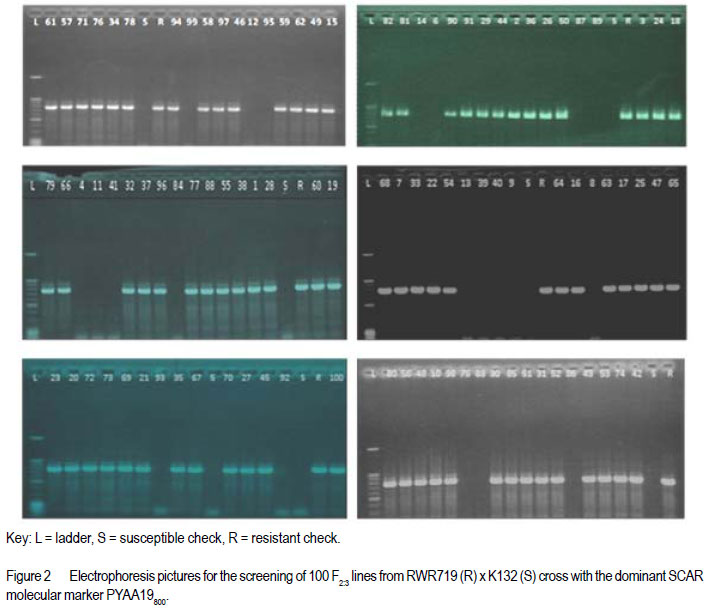
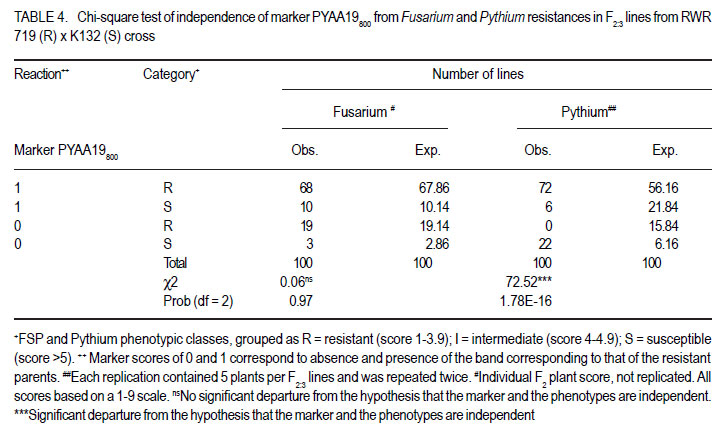
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 31-39 Evaluating genetic association between fusarium and pythium root rots resistances in the bean genotype RWR 719 P. O. Ongom, S.T. Nkalubo1, P.T. Gibson2, C.M. Mukankusi3 and P.R. Rubaihayo College of Agricultural and Environmental Sciences, Makerere University, P. O. Box 7062, Kampala, Uganda Code Number: cs12011 ABSTRACT Resistance to Fusarium root rot (Fusarium solani f.s.p phaseoli) has been reported in common bean (Phaseolus vulgaris L.) sources and is usually associated with Pythium root rot resistance. Pythium root rot (Pythium ultimum var ultimum) resistance is controlled by a single dominant gene, marked by a SCAR marker PYAA19800 . It remains unclear whether the inheritance to resistance of these two bean root rots is genetically independent. We evaluated the association of Fusarium root rot resistance with the Pythium root rots resistance gene and/or the molecular marker PYAA19800 in genotype RWR 719. Two populations; F2 and F2:3 lines, generated from RWR 719 (resistant) × K132 (susceptible) were respectively screened with Fusarium solani and Pythium ultimum isolates, and root damages were scored based on the CIAT 1 – 9 scale. Additionally, the F2:3 lines were screened with PYAA19800. The F2 segregation ratio deviated from a single gene model for reaction to Fusarium solani. The F2:3 lines fit the model for a single dominant gene that confers resistance to Pythium ultimum. Fusarium solani and Pythium ultimum resistances were inherited independently. There was lack of association between PYAA19800 and Fusarium solani resistance, but the PYAA19800 was strongly associated with Pythium ultimum resistance. This contradicts the assertion of linkage of the two resistances that was deduced based on the joint occurrence of both resistances in the available donor genotypes. Key Words: Bean root rots resistance, Fusarium solani, molecular marker, Phaseolus vulgaris, Pythium ultimum RÉSUMÉ La résistance à la porriture racinaire par suite du Fusarium solani f.s.p Phaseoli a été signalée dans les sources du haricot commun (Phaseolus vulgaris L.) et communément associée à la résistance de la pourriture racinaire due au Pythium. La résistance à la pourriture racinaire due au pithium (Pythium ultimum var ultimum) est controlée par un unique gène dominant marqué par un marquer SCAR PYAA19800. Il demeure incertain si l’héritage de la résistance à ces deux types de pourriture racinaire du haricot est génétiquement indépendant. Nous avons évalué l’association des gènes de résistance à la pourriture racinaire due au Fusarium solani et au Pythium ultimum et/ou le marqueur moléculaire PYAA19800 dans le génotype RWR 719. Deux populations, F2 et F2:3 issues du croisement entre 719 (résistant) × K132 (sensible) ont été respectivement testées avec des isolats de Fusarium solani et Pythium ultimum, et les dégâts étaient mesurés sur base de l’échelle de cotation de 1 à 9 du CIAT. Additionnellement, F2:3 étaient testées avec PYAA19800. Le rapport de ségrégation de F2 a dévié du modèle unique pour la réaction au Fusarium solani. Les lignées F2:3 se sont adaptées au modèle de l’unique gène dominant qui confère la résistance au Pythium ultimum. Les résistances au Fusarium solani et au Pythium ultimum étaient indépendamment héritées. Il n’y avait aucune association entre PYAA 19800 et la résistance au Fusarium solani, mais le PYAA19800 était un gène unique dominant avec résistance au Pythium ultimum. Ceci contrédit l’assertion du lien de ces deux résistances déduit sur base de l’occurrence combinée de toutes les deux resistances dans les génotypes donneurs disponibles. Mots Clés: Bean root rots resistance, Fusarium solani, molecular marker, Phaseolus vulgaris, Pythium ultimum INTRODUCTION Root rot of bean (Phaesolus vulgaris L.) is a major constraint to bean production in Uganda, causing total crop failures in some seasons (Spence, 2003; Namayanja et al., 2003; Kakaire, 2008). Bean root rot is caused by a complex of soil-borne pathogens, the most important ones being Fusarium solani f. sp. Phaseoli and Pythium ultimum var. ultimum (Rusuku et al 1997; Spence, 2003; Tusiime, 2003). Root rot symptoms due to Fusarium sp. and Pythium sp. include poor seedling establishment, damping-off, uneven growth, chlorosis, premature defoliation, death of severely infected plants, foliar blight or pod rot, and lower yield (Abawi and Pastor-Corrales, 1990; Román-Avilés et al., 2003; Abwai et al., 2006). The two pathogens occur concurrently in farmer’s fields (Tusiime, 2003). Management of bean root rot diseases is complicated due to the involvement of multiple soil-borne pathogens, and by the fact that the pathogens produce oospores that are highly persistent in the soil for several years (Abwai et al., 2006). Several management practices are used to control Fusarium and Pythium root rots; seed or soil treatments with selective pesticides, crop rotations, cover crops, seedbed preparations, among other measures, may improve yield or reduce root rot severity (Abwai et al., 1985; CIAT, 2003). These measures have not been consistently economical for the resource poor farmers (CIAT, 2003). Exploiting host plant resistance is considered as the most effective way of managing bean root rots for small-scale farmers (CIAT, 2003; Otsyula et al., 2005). Resistance to Fusarium solani is complex and is conditioned by two or more genes (Schneider et al., 2001; Romans-Aviles, 2005; Mukankusi et al., 2011), whereas, Pythium ultimum resistance is controlled by a single dominant gene, marked by a dominant SCAR marker-PYAA19800 (Otsyula et al., 2003; Mahuku et al., 2005; Otsyula, 2010). The presence of joint resistance to both Fusarium and Pythium root rots have been observed in several resistance sources (Spence, 2003; Tusiime, 2003; Mukankusi, 2008), and it is thought that genes conferring resistance to Pythium root rot may be closely linked or even the same loci (pleiotropic) as those conferring resistance to Fusarium root rot. Quantitative Trait Loci (QTLs) linked to Fusarium solani resistance have been mapped on the same chromosome as that on which gene for resistance to Pythium ultimum has been found (Romàn-Avilès and Kelly, 2005 and Mahuku et al., 2005), however, it is not clearly known whether the two traits are associated. Dickson and Boettger (1977) using one resistant source (Cornell 2114-12) observed low correlations between resistances to Fusarium and Pythium root rots. They however, used mixed cultures of both pathogens which could have affected the evaluation of co-segregation. It is known that when the two pathogens are together, they exhibit synergistic interactions, exaggerating disease severity (Spence, 2003), and since they show similar symptoms, it is difficult to score the severity of both pathogens on the same plant. In this study pure cultures and a molecular marker technique to evaluate any possible genetic link between resistances to Fusarium solani and Pythium ultimum root rots in the genotype RWR 719, which is resistant to both pathogens were used. MATERIALS AND METHODS Genetic plant materials. The population was developed by crossing RWR 719 (R) with K132 (S) in 2008 at National Crop Resources Research Institute Namulonge (NaCRRI). During 2009, the obtained F1 seeds were planted in pots in the screenhouse at NaCRRI and were bulk harvested. The F2 population was evaluated for resistance to Fusarium root rots at National Laboratories Research Institute at Kawanda (NALRI), and the F2 plants were rescued after 21 days, transplanted in pots, and harvested individually. This generated 100 F2:3 lines that were screened with both Pythium ultimum isolate and the Pythium molecular marker, PYAA19800 . Evaluation of populations using Fusarium solani isolate. The most pathogenic isolate of F. Solani f.sp. phaseoli (FSP-3) that had been identified by Mukankusi (2008) and preserved at NALRI was used. To rejuvenate the FSP-3 isolate, pure colonies stored on potato dextrose agar (PDA) slants at 5OC were grown on fresh PDA plates for a period of 21 days and then multiplied on sterilised sorghum seed. The inoculum was prepared following the method described by Mukankusi (2008). One bottle of inoculum (500ml capacity) was added to pre-sterilised sandy-clayloam soil in each tray (0.74 x 0.42 x 0.115m3). A susceptible line, K132, was then planted in each tray for up to 28 days, when it was uprooted. This was done to test the pathogenicity of the FSP-3 isolate and to increase disease pressure in the soil before planting the test materials. To ensure uniform levels of inoculum in all trays, soil was removed from all the trays, remixed together and redistributed equally into the trays prior to planting the test populations. The test populations (F2 seeds) were then planted in four wooden trays containing mature FSP-3 inoculum. Each tray was planted with 8 rows of the test material and 2 rows of each of the resistant (RWR 719) and susceptible (K132) parents. The trays were placed on a raised wire mess in the screenhouse and watered four times a week (Mukankusi, 2008). Reactions to disease were assessed 28 days after planting by washing the below- ground parts of the plant (hypocotyls and roots) under running tap water, and scoring the reaction according to the C1AT 1-9 scale (Abawi and Pastor-Corrales,1990). In this scale; 1 = no visible symptoms 3 = light discolouration either without necrotic lesions or with approximately 10% of the hypocotyl and root tissue covered with lesions; 5 = approximately 25% of the hypocotyl and root tissue covered with lesions though the tissues remain firm, with some deterioration of the root system; 7 = approximately 50% of the hypocotyls and root tissues covered with lesions combined with considerable softening, rotting, and reduction of root system; and 9 = approximately 75% or more of the hypocotyls and root tissues affected with advanced stages of rotting, combined with severe reduction in the root system. Evaluation of populations using the Pythium ultimum isolate. The pathogenic isolate of Pythium ultimum var.ultimum (MS 61), which had previously been isolated and preserved at CIAT-Kawanda (Mukalazi et al., 2001), was used for this study. A total of 10 seeds of each of 100 F2:3 families from RWR x K132 cross were planted in wooden trays containing soil infested with the MS 61 isolate. A tray consisted of two checks and 20 F2:3 families; each planted in short rows of five plants per family. The experiment was laid out in a randomised complete block design with two replications of five trays per replication. Germinated seedlings were watered twice a day for three weeks to provide favorable conditions for infection, establishment and development of the fungus (Mukankusi, 2008). Individual seedlings were uprooted after 3 weeks, washed in tap water, and scored according to the CIAT 19 scale (Van Shcoonhoven and Pastor-Corrales, 1987). Plants with limited or no symptoms (score 1-3) were rated as being resistant, and the rest of the plants as susceptible. The average score for each of the 100 F2;3 families was obtained, and the class frequencies were tested for goodness of fit to the theoretical ratios, using the chi-square test. The correlation between the average scores for Pythium root rot and the Fusarium root rot phenotypic scores was evaluated. Evaluation of populations with thePythium SCAR marker PYAA19800 . This experiment was conducted to investigate the association of the Pythium molecular marker (PYAA19800) with F. solani and P. ultimum resistances. Young trifoliate leaves were sampled from six plants for each of the 100 F2:3 families described, and the extracted DNA from the plants in the same family was later bulked since each family was derived from a single F2 plant. Six plants were sampled in order to have a high probability of capturing at least one allele from the resistant parent in each segregating family. One half of the F2:3 families were expected to segregate (1RR: 2Rr: 1rr). Total genomic DNA was extracted using the procedure described by Mahuku (2004). Extracted DNA was quantified with a DQ200 fluorometer following the manufacturer’s instructions, and DNA from each sample was then diluted based on concentration of each sample to 20 or 80 ng.ml-1 for polymerase chain reactions (PCR). The PCR master mix was constituted to consist of 0,2 mM of dNTPs, 2mM MgCl2, 1µ/25µl of Taq Polymerase, 1X PCR Buffer and 0,4 µM of each primer. The sequences of the primers used were: 5’ -TTA GGC ATG TTAATT CAC GTT GG-3’ and 5’-TGA GGC GTG TAA GGT CAG AG- 3’ (CIAT, 2008). The 25µl-reaction PCR volume was subjected to 34 amplification cycles in a BIO RAD MyCycler thermal cycler. The amplification consisted of one cycle at 94°C for 5 minutes, followed by 34 cycles of: denaturation at 94ºC for 40 seconds, annealing at 63°C for 40 seconds, and extension at 72°C for 40 seconds for 34 cycles, followed by a final extension for 7 minutes at 72°C and a holding temperature of 4°C. The amplified products were separated according to size, using 1.2% agarose gel in a 0.5X TBE buffer at a voltage of 100V for 45 minutes, and then stained with ethidium bromide (0.5µg/ml) to be visualised under ultraviolet light and photographed before scoring. Bands were scored as 1 or 0 for presence or absence of a specific DNA fragment. This data was compared with FSP phenotypic scores to detect a possible association. Association analyses. A genetic linkage test was performed based on chi-square test of independence, using phenotypic classes (R, S) for Fusarium and Pythium root rots in order to assess the association of resistances. This result was supported by running a correlation analysis in Genstat (Genstat, 2010). To assess the association of the PYAA19800 marker locus with the Fusarium solani phenotypic class (R or S), a chi-square test of independence was performed. A regression analysis of PYAA19800 against Fusarium root rot score was performed in Genstat to estimate the magnitude of the marker effect. RESULTS When F2 populations of RWR 719 x K132 cross were inoculated with Fusarium solani isolate, the 100 F2 plants segregated into 87 resistant : 13 susceptible. Segregation ratio for the F2 population deviated from a 3: 1 gene model (χ2 = 7.68, P = 0.006) (Table 1). For the 100 F2:3 lines of the same cross, the segregation ratio was 72 resistant : 28 susceptible for reaction to Pythium ultimum which fit the 3:1 model for a single dominant gene (χ2 = 0.48, P = 0.49) (Table 1). The chi-square test of independence for resistance to both Fusarium Solani and Pythium ultimum in RWR 719 × K132 cross showed no significant relationship between the two pathogens (χ2 = 4.96, P = 0.29) (Table 2). These results were further supported by a correlation analysis that displayed no significant association between the two traits (R2 = 0.007, P = 0.39, Fig. 1). When 100 F2:3 lines were screened with PYAA19800 (a dominant SCAR marker), 78 lines produced bands that indicated the presence of the Pythium resistance gene and 22 lines showed no band, indicating absence of the gene (Fig. 2). The marker scores fit a 3:1 ratio (χ2 = 0.48, P = 0.49) for a dominant marker indicating normal segregation in a simple Mendelian ratio (Table 3). Results of the test of independence showed no significant association of Fusarium root rot resistance with the Pythium marker PYAA19800 (χ2 = 0.06, P = 72.5), although the marker was strongly associated with Pythium root rot resistance (χ2 = 0.97, P = 1.78E-16) (Table 4). The results from the test of independence were very consistent with the regression analysis (Table 5). DISCUSSION The current study revealed that Pythium root rot resistance was conditioned by a single dominant gene. Also, based on earlier studies, Hassan et al. (1971), Boomstra and Bliss (1977), Faria (1983) Schneider et al. (2001), Romans-Aviles and Kelly (2005), Mukankusi (2008) among others reported that two or more genes govern Fusarium resistance which closely agree with the present finding. Consistent with Buruchara et al. (2001, Unpubl.), Otsyuala et al. (2003), and Otsyuala (2010). Results from the study, are at variance with previous findings (Tusiime, 2003; Spence, 2003; Mukankusi, 2008) indicated that Fusarium and Pythium root rots resistances in RWR719 were independently inherited. It is probable that the joint resistance in genotype RWR 719 was a result of field selection of materials with both Fusarium and Pythium root rots resistances since the two pathogens frequently occur together in the field. Genetic independence of the inheritance of resistance for the two bean roots rots is in line with findings by Dickson and Boettger (1977) who observed low correlations between the two traits. Additionally, marker analysis revealed lack of association between Fusarium solani resistance and the Pythium marker PYAA19800 . As expected, Pythium marker PYAA19800 was closely associated with Pythium ultimum resistance in RWR 719, explaining over 77% of phenotypic variability, and confirming its suitability for use is breeding for P. ultimim resistance. Mahuku et al. (2005) had reported an association of PYAA19800 with Pythium ultimum resistance in RWR 719, MLB 49-89A and AND 1062, located 1.5cM from the resistance gene. ACKNOWLEDGEMENT This research was supported by the Alliance for a Green Revolution in Africa/Program for African Seed Systems (AGRA/PASS) project. The authors thank Dr. Richard Edema for his support in implementation of the study, NaCRRI-Namulonge where populations were developed, and CIAT-Uganda who provided the isolates used in the study. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12011t3.jpg] [cs12011t1.jpg] [cs12011f1.jpg] [cs12011t5.jpg] [cs12011t4.jpg] [cs12011t2.jpg] [cs12011f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}