 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
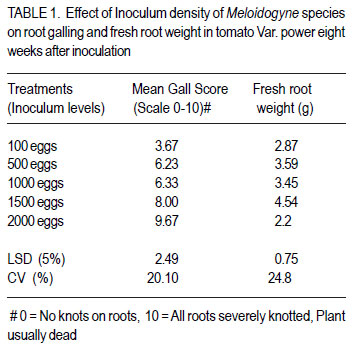
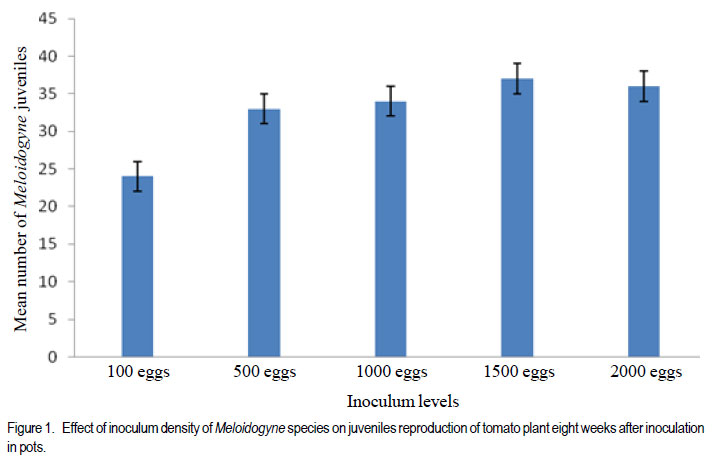
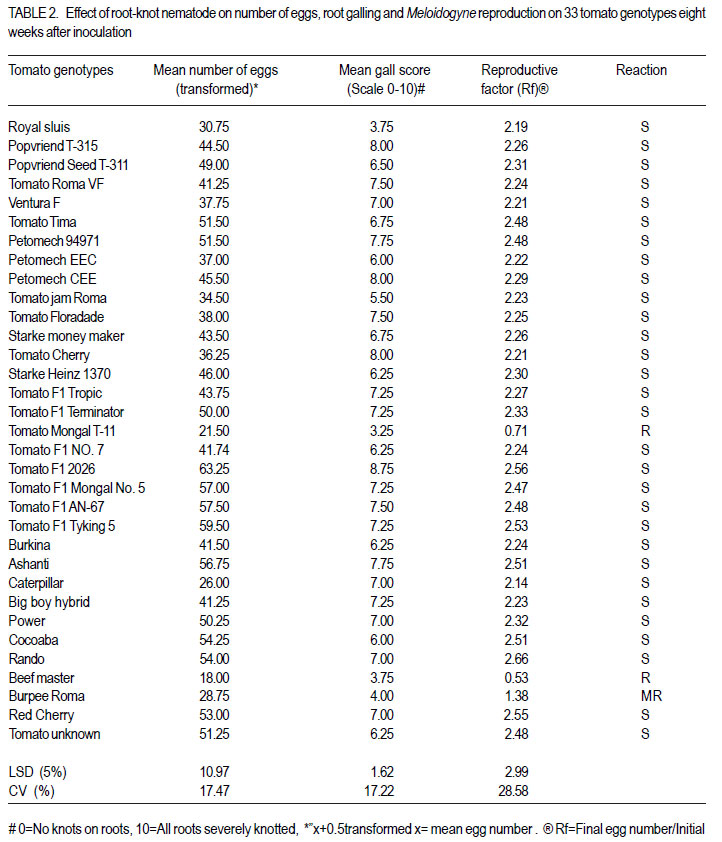
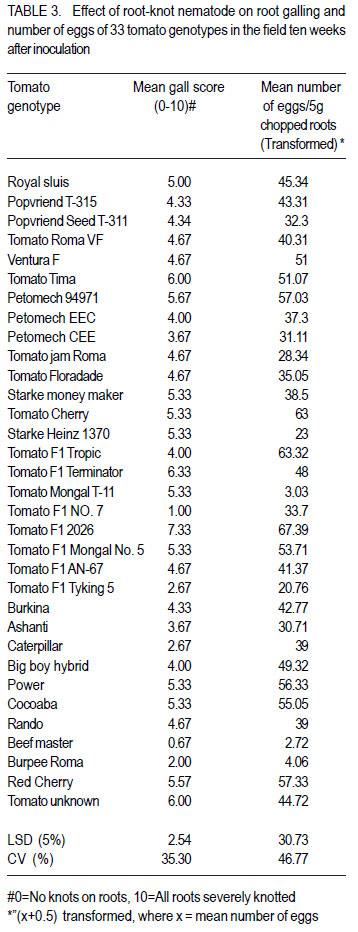
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 41-49 Evaluation of tomato genotypes for resistance to root-knot nematodes F. Jaiteh, C. Kwoseh1 and R. Akromah1 National Agricultural Research Institute (NARI) – PMB 526, Serrekunda, The Gambia Code Number: cs12012 ABSTRACT Tomato (Solanum lycopersicum) is one of the most popular vegetable crops worldwide, owing to its high nutritive value and diversified use. Tomato production in Ghana is threatened by plant parasitic nematodes, especially the root knot nematodes (Meloidogyne spp.), which are responsible for huge economic yield losses. The losses, however, can be averted through use of resistant varieties. A study was conducted to evaluate tomato genotypes reaction to root-knot nematodes. Five inoculum levels, 100, 500, 1000, 1500 and 2000 root-knot nematode eggs per 1.6 litres of soil per pot were inoculated on tomato seedlings. A total of 33 genotypes were screened for resistance to Meloidogyne spp. Nine tomato genotypes were collected from Burkina Faso, eight from Vietnam, seven from the United States of America, three genotypes from South Africa, and six from local agro-dealers and farmers in Ghana. The genotypes were screened in pots and the field for host resistance to Meloidogyne spp. The 1500 eggs per plant inoculation level resulted in the highest number of eggs, juveniles and fresh root weight. Among the 33 genotypes screened, Tomato Mongal T-11 and Tomato Beef Master were found to be highly resistant to Meloidogyne spp. and also recorded the lowest reproductive factors of 0.71 and 0.53, respectively. Key Words: Host resistance, Meloidogyne sp., Solanum lycopersicum RÉSUMÉ La tomate (Solanum lycopersicum) est une des plus populaires légumes au monde suite à sa valeur plus nutritive et utilisation diversifiée. Par ailleurs, sa production est limitée par des nématodes parasites des plantes, spécialement les galles racinaires (Meloidogyne spp.) et sont responsables de lourdes pertes économiques. Ces pertes, par ailleurs, peuvent être évitées par l’utilisation des variétés résistantes. Une étude était conduite pour évaluer la réaction des génotypes de tomates aux galles racinaires des nématodes. Cinq niveaux d’inoculum à savoir 100, 500, 1000, 1500 et 2000 galles d’oeufs de nématodes par 1.6 litres de sol par pot étaient inoculés sur les plantules de tomates. Un total de 33 génotypes étaient résistants au Meloidogyne spp. Neuf génotypes de tomates étaient collectés du Burkina Faso, huit du Vietnam, sept des Etats Unis d’Amérique, trois génotypes d’Afrique du Sud et six provenant des vendeurs et fermiers locaux du Ghana. Les génotypes étaient testés en pots et en champ pour résistance de l’ hôte au Meloidogyne spp. Les 1500 oeufs par niveau d’inoculation de plants ont induit un nombre le plus élevé d’oeufs, raciness juveniles et le poids des racines fraîches. Parmi les 33 génotypes testés, Tomato Mongal T-11 et Tomato Beef Master ont été trouvés les plus résistantes au Meloidogyne spp. et avaient aussi enregistré les facteurs les moins reproductifs de 0.71 and 0.53, repectivement. Mots Cles: Résistance de l’hôte, Meloidogyne sp., Solanum lycopersicum INTRODUCTION Tomato (Solanum lycopersicum L.) is consumed nearly in every household in Ghana and it accounts for 38% of the total vegetable expenditure (Wolff, 1999). Beside being tasty, tomatoes also contribute to a healthy diet being a source of vitamins A, B and C and also containing good amounts of potassium, iron, and phosphorus (Wener, 2000). According to Horna et al. (2006), in Ghana during the dry season, local production is not able to meet the domestic high demand and tomatoes are often imported, mainly from Burkina Faso. This can be attributed to several reasons, and most important among these is the vulnerability of tomato crop to various diseases including fungal, viral, bacterial and nematode diseases (Sasser et al., 1983). Unlike other pathogens, nematodes are more challenging to control because they live in the soil and cannot be easily seen by farmers (Mai, 1977). They are only noticed when the population is widespread and yield is very low. Root knot nematodes are one of the major pathogens of tomatoes worldwide and limit fruit production (Sikora and Fernandez, 2005). An estimated amount of US$500 million is spent on root-knot nematode control globally (Keren-Zur et al., 2000). Yield losses of between 73 - 100% were reported in Northern Ghana due to root-knot nematode (Hemeng, 1981). They cause serious damage to tomatoes, impacting both the quantity and quality of marketable yields. In addition to the direct losses, root-knot nematodes also interact with other plant pathogens such as fusarium, resulting in increased damage (Peacook, 1957). A range of strategies are used to control rootknot nematodes, and these include the use of nematicides, organic manure amendment and resistant cultivars. In Ghana, most farmers use synthetic chemicals indiscriminately to control nematodes and associated infections to tomato. However, chemical nematicides have disadvantages in being expensive and extremely toxic to man and animals when not applied at the right time, in the right way and in the right dosage. Their use is also curtailed because of their threat to groundwater as well as long waiting periods between use, harvesting and marketing of crops (Bridge, 1996). From the foregoing, genetic resistance in tomato against root-knot nematodes will be most efficient in reducing nematode population densities and, thereby, reducing the need for pesticide application. Resistance cultivars also do not require additional cost input to the farmer. Most importantly, they do not leave toxic residues on plants. These facts support the need to identify more genotypes for resistance against root-knot nematodes in tomato. The objective of this study was to determine the best egg inoculum level for screening tomatoes for resistance to root-knot nematodes to avoid misclassification and to evaluate tomato genotypes for their reaction to root-knot nematode. MATERIALS AND METHODS The study was carried out at the Plant Pathology laboratory and Plant house of the Department of Crop and Soil Sciences, Kwame Nkrumah University of Science Technology (KNUST), and the agricultural research experimental field at KNUST. Three different experiments were conducted in this study. Sources of tomato genotypes and root-knot nematodes inoculum. A total of 33 genotypes of tomato were evaluated for resistance to Meloidogyne species. Nine of the genotypes were collected from Burkina Faso, three from South Africa, seven from the United States of America, eight from Vietnam, and six from agrodealers and farmers in Ghana. The root-knot nematode inoculum used was obtained from heavily infested tomato roots collected from vegetable farms around KNUST, Kumasi. Extraction and counting of root-knot nematodes eggs and juveniles. A modified Baermann tray (Whitehead and Hemming, 1965) was used to extract nematode juveniles from roots. Eggs were extracted using modified Hussey and Barker (1973) method. Counting was done with the aid of a stereo microscope, counting tray and tally counter. Soil preparation and sterilisation. Soil for pot experiments was sterilised using the barrel steam sterilisation method. Black soil was mixed with river sand at 3:1 ratio (v/v) and sterilised for 3 hr at 100 o C with fuel wood as the source of heat. Black soil used for the tomato nursery was sterilised using sterilisation method above. Land preparation and harvesting of the tomato genotypes. The land was slashed, ploughed, hallowed and ridges made at 1m between rows and 20 cm within rows. The experiment design was a randomised completely block design with three replications. The test plants were harvested three months after transplanting. To ensure easy removal of the plants from the soil, the sides of the ridges were dug and the plants were carefully lifted from the ground. The soil was then removed from the roots by gently shaking the plants. The roots were cleaned as described above. Nursing of tomato seeds and transplanting of seedlings. The different tomato seeds collected were nursed separately in plastic seed trays containing the sterilised black soil. Three weeks after germination, the most uniform seedlings were transplanted to pots for their reactions to root knot nematodes. Each pot of 2 L size was filled with 1.6 L of the sterilised black soil-river sand mix. One tomato seedling per pot was used. Application of different Meloidogyne inoculum levels to potted tomato seedlings The potted seedlings were inoculated with 100, 500, 1000, 1500 and 2000 eggs per pot two weeks after transplanting. Three wells were made in a triangular form 2 cm from each plant. The egg suspension of each level was hormogenised by blowing air through a pipette and then dispensed into the holes. The control was without inoculation (0 eggs). Complete randomised design with four replicates was used. Plants were watered every 24 hours with 500 ml of tap-water. Harvesting of tomato plants. The test plants were harvested eight weeks after inoculation. To ensure easy removal of the plants from the soil, the sides of the plastic pots were pressed to loosen the soil. The soil was then removed from the roots by gently shaking the plants. The roots of the harvested tomato genotypes were each washed separately and dabbed dry with tissue paper. Assessment of the tomato plants for root knot nematode infection. The roots of the harvested tomato plants were each washed separately and dabbed dry with tissue paper. Galling was scored on scale of 0-10 rating chart by Bridge and Page (1980). Fresh weight of roots of each entry in the screenhouse was measured using an electronic balance. Galling and Fresh roots weights were also recorded for the field experiment. Reproduction factor (Rf) which is the final population divided by the initial population was calculated. A Rf greater than 1, connotes reproduction, while Rf of less than 1, implies no reproduction. Data collected. Data were collected from the following variables: Plant height. Plant height was measured as the length from the top of the shoot to the base of each plant using a meter ruler. Stem diameter. Stem girth was measured using a thread, and a ruler. A thread was stretched around the stem of each plant and subsequently place on a ruler to take the reading. Root gall score. Galling was scored on the scale of 0-10, using a rating chart by Bridge and Page (1980). Fresh shoot and root weight. After eight weeks of inoculation, the test plants were uprooted gently. In order to ensure easy removal of the plants from the soil, the sides of the soil was pressed to loosen the soil. The soil was then removed from the roots by gently shaking the plants. Fresh shoot and roots weight of each entry was measured using an electronic balance by placing the plant on the scale and taking the readings. Number of root-knot nematodes juveniles. Extraction of nematode eggs was done by using modified Hussey and Barker (1973) method. Root knot nematodes–infested tomato roots were washed, dabbed dry and then cut into pieces with a pair of scissors. A reasonable quantity of the chopped roots was placed in a big jam bottle and 0.5% sodium hypochlorite (NaOCl) solution was added to cover the roots and then covered. The content of the bottle was agitated vigorously for four minutes. The chopped root-NaoCl suspension was then collected and rinsed with tap water on 200 ìm-pore mesh sieve over 500 ìmpore mesh sieve and rinsed with tap water was added to obtain the actual egg-water suspension. Root knot nematode eggs were counted using a counting tray with the aid of a stereo microscope. Counting was done three times per entry. Root-knot nematodes. Juveniles were extracted from infested tomato roots using modified Baermann tray method (Whitehead and Hemming, 1965). The roots were chopped with a pair of scissors and 5 g weight of each entry was placed in a plastic sieve lined with a ply- two tissue paper placed in a plastic plate. Tap-water was poured gently into the plastic plate in which the sieve was placed until the tissue became moist. The set up was left for 48hr and the plates were then poured separately into beakers and left overnight for the juveniles to settle. Each nematode water suspension was separately topped with tap water to 30 ml for standardisation. Each suspension was homogenised by blowing air through with a pipette. Counting was done three times to obtain the mean number of juveniles. Experimental designs and data analyses. The screenhouse study was set up in completely randomised design with four replicates; while a randomised complete block design with three replicates was used for the field experiment. Data collected were analysed using the GenStat statistical package (Discovery edition 3). Where the ANOVA indicated significant treatment differences, the Least Significant Difference (LSD) at 5% was used for comparing means. All count data were transformed using square root transformation of “(x+0.5), where x was the mean count. RESULTS AND DISCUSSION The best inoculum level for screening tomato for root-knot nematodes resistance. Generally, when inoculum level increased, mean gall score also increased (Table 1). The roots of tomato plants inoculated with 2000 eggs/plant had the largest gall infestation of 9.67 as compared with all inoculum levels. The 100 eggs inoculum density/tomato plant scored the least gall index of 3.67 (Table 1). There was no diference (P<0.05) between the 2000 eggs/tomato plant and 1500 eggs/tomato plant, but the former was significantly different (P<0.05) from all other treatments. The 1500 eggs/tomato plant recorded fresh root weight of 4.54 g, which was significantly more than that of for the 100 eggs/tomato plants which had the least fresh weight of 2.87 g. The fresh root weight of the 1500 eggs/tomato plant was significantly different (P<0.05) from all the other treatments (Table 1). These obsevations are in accordance with reports by Nadary et al. (2006) and Chindo et al. (2006) that as initial inoculum level of M. incognita populations increased, populations that had high infection incidence and reproduction rates induced greater root galling than did other populations. Wonang and Akueshi (1990) studied the relationship between population densities of M. incognita and crop yield in tomato and found that there was greater increase in the fresh weight of tomato roots with increase in the inoculum density. This is inline with the observation in Table 1. The number of Meloidogyne juveniles ranged from 24 to 47 for 100 eggs/ tomato plant and 1500 eggs/tomato plant, respectively. There was significant differences (P<0.05) between the treatments except between the 500 eggs/tomato plant and1000 eggs/tomato plant and 1000 eggs/ tomato plant and 2000 eggs/tomato plant (Fig. 1). Results of the study revealed that increasing initial population of Meloidogyne spp. in the soil had a direct effect of increasing juvenile population in the tomato root. However, the 2000 eggs/tomato plant root system could not support increased juvenile population probably because of limited living tissue resulting in decreasing population. This relationship reflects the density dependent effect of increased competition for feeding sites and food reserves at high initial densities of nematodes (Riaz et al., 2000). This observation is also in agreement with the report by El-Sheriff et al. (2007) who studied the effect of fifteen population densities of M. incognita race 1 ranging from zero to 5000 eggs on yield of tomato, and found that maximum number of nematode juveniles were recorded at moderate population density (1000 to1500 eggs); and at very high population densities the reproduction potentials of root-knot nematodes on the plant declined because the population in the root system reached its peak and could not support further reproduction. Niblack et al. (1986) demonstrated that at moderate to high initial population densities, of root-knot nematodes reached their maximum levels on a susceptible cultivar. Whereas on partially resistant cultivars had less damage by the nematodes, the population densities were still increasing. Reaction of tomato genotypes to root-knot nematodes (Meloidogyne species) in pots. The mean root-knot nematode eggs recovered from the roots of the 33 tomato genotypes ranged from 18.00 to 63.25 for Beef master and Tomato F1 2026, respectively. There was no difference (P>0.05) between Beef master, tomato mongal T11 and Burpee roma (Table 2). Tomato F1 2026 also recorded the largest gall infestation score compared with the other genotypes. According to the gall score rating used for host response, nematode mean gall score was lowest (3.25) in Tomato mongal T-11 followed by Beef master (3.75) and then Burpee roma(4.00) as compared to rest of the genotypes (Table 2). All genotypes showed great variation in response or reaction to root-knot nematodes from resistant to susceptible (Table 2).Out of the 33 tomato genotypes screened, two namely, Mongal T-11 and Beef Master were found to be resistant, while Burpee roma was moderately resistant(MS), and the rest susceptible (S) to Meloidogyne species (Table 2). Karssen and Moens (2006) reported that highly susceptible host plants allowed the juveniles to enter the roots, reached maturity and produced many eggs, while the resistant plants suppressed their development and thus, do not allow reproduction. According to Cousins and Walker (1998), root-knot nematode eggs developed poorly on resistant accession compared to susceptible accession. Also, the authors reported that quantity of eggs reflected on the number of nematodes that reached reproductive maturity, and therefore provided one measure of resistance. Fassuliotis (1979) reported that because galling occurs in most susceptible plants infected with root-knot nematodes, this can often be the sole measurement of resistance during screening. Field assessment of 33 tomato genotypes. More eggs were recovered from the roots of Tomato F1 2026, Tomato cherry and Red cherry, as compared with Tomato Mongal T-11, Tomato Beef master and Burpee Roma (Table 3). Roots of susceptible genotypes were found to be more favourable to root-knot nematode galling (Table 3). Therefore, more eggs were identified on susceptible genotypes roots compared with the less susceptible and/or resistant genotypes. Root galling on tomato varied with different genotypes. Lower gall score was recorded on the roots of Tomato Mongal T-11 as compared to Tomato F1 2026. The primary symptom of root-knot nematode infection is the formation of typical root galls on the roots of susceptible genotypes. The presence or absence of root galls on tomato plants indicates whether a variety is resistant or susceptible to root-knot nematodes. However, significant differences in the number of galls present on roots indicate different levels of susceptibility. Khan (1994) reported that nematode resistance in host plants was manifested by reduced rates of nematode reproduction, egg masses and consequently, low nematode population densities than that of a susceptible one. Thus, the above observation is in accordance with the report by Khan (1994) that the development of galls on plant roots increased significantly on the susceptible genotypes compared with resistant genotypes thereby affecting plant performance. Mai and Abawi (1987) reported that intensive galling significantly reduced root efficiency translanting into yield reduction. ACKNOWLEDGEMENT The authors thank the National Agricultural Research Institute (NARI) –The Gambia and Strengthening Capacity for Agricultural Research and Development in Africa (SCARDA) for funding this research. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12012t2.jpg] [cs12012t1.jpg] [cs12012f1.jpg] [cs12012t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}