 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
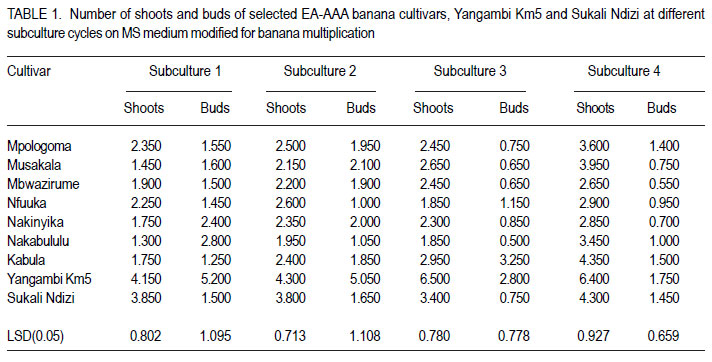
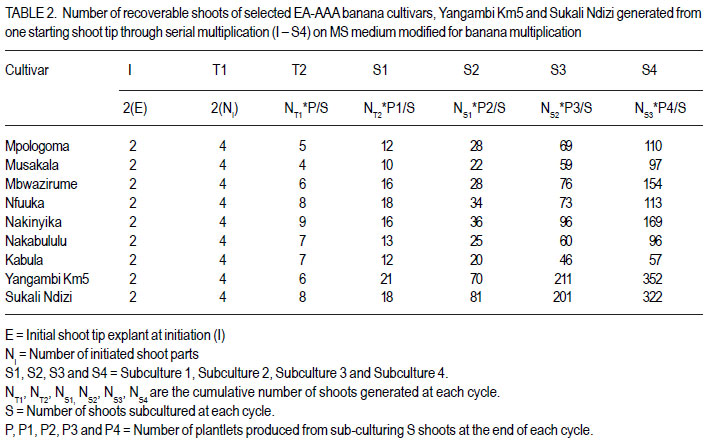
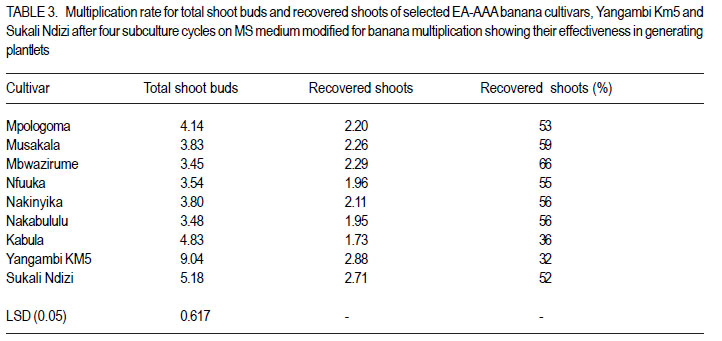
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 67-76 Proliferation and shoot recovery among the East African highland banana K. Sadik, G. Arinaitwe1, J.M. Ssebuliba, P. Gibson, C. Lugolobi and S.B. Mukasa College of Agricultural and Environmental Sciences, Makerere University, P. O. Box 7062 Kampala, Uganda Code Number: cs12015 ABSTRACT Production of East African highland banana (EA-AAA banana) (Musa spp.) is limited by scarcity of planting materials, attributable to their low natural proliferation ability. Under natural field conditions, the EA-AAA bananas greatly differ in suckering ability. In vitro micropropagation has been adopted as an alternative means for production of banana planting materials. In this study, the in vitro proliferation potential of seven EA-AAA banana cultivars, with different suckering ability was determined on Murashige and Skoog (MS) medium, to enhance development of micropropagation protocols for their multiplication. Commonly cultivated non EA-AAA banana cultivars were used to compare proliferation of the seven EA-AAA cultivars. There was a wide variation in the number and morphology of shoots and buds produced by the different cultivars. The EA-AAA banana cultivars produced 3-4 new shoots in each subculture cycle, and 57-169 recoverable shoots from one starting shoot-tip explant in 18 weeks. Non-EA-AAA banana cultivars, namely Sukali Ndizi (AAB) and Yangambi Km5 (AAA), showed higher proliferation levels, 5 and 9 shoots, from each subculture cycle and 322 and 352 recoverable shoots, respectively. The EA-AAA banana cultivars showed higher efficiency to produce recoverable shoots from shoot buds (53 - 66% except for cv. Kabula at 36%) compared to Sukali Ndizi (52%) and Yangambi Km5 (32%). The study demonstrates the potential of in vitro approach for production of banana planting materials. In vitro proliferation ability and in particualr efficiency to produce recoverable shoots of the different EA-AAA banana cultivars could be improved by varying the culture conditions during the subsequent subculture cycles. Key Words: EA-AAA banana, In vitro micropropagation, Musa spp. shoot buds RÉSUMÉ La production de la banane (EA-AAA banana) (Musa spp.) dans les montagnes de l’Afrique de l’Est est limitée par le manque du matériel de plantation suite à leur base capacité de prolifération. En conditions naturelles au champs, les bananes EA-AAA different considérablement en leur capacité de succion. La propagation in vitro a été adoptée comme moyen altérnatif pour la production du matériel de plantation de la banana. Dans cette étude, le potentiel de prolifération in vitro de sept cultivars de banana EA-AAA de capacité de succion différente, était déterminé sur les media de Murashige et Skoog (MS), afin d’améliorer le développement de protocoles de micropropagation pour leur multiplication. Des cultivars communément cultivés autre que la banana EA-AAA étaient utilisés pour faire la comparaison avec la proliferation des sept cultivars EA-AAA. Il y’avait eu une large variation dans le nombre et la morphologie des pousses et bourgeons de différents cultivars. Les cultivars de bananes EA-AAA ont produit 3-4 nouvelles pousses dans chaque cycle de sous culture et 57-169 pousses recouvrables d’un explant de pousse dans 18 semaines. Les cultivars de bananes non-EA-AAA nomément Sukali Ndizi (AAB) and Yangambi Km5 (AAA), ont montré de niveaux élevés de proliferation, 5 et 9 pousses de chaque cycle de sous culture et 322 et 352 pousses recouvrables, respectivement. Les cultivars de bananes EA-AAA ont montré un niveau d’efficacité élevé quant à la production de de pousses recouvrables à partir de bourgeons (3566% excepté pour cv. Kabula à 36%) en comparaison avec Sukali Ndizi (52% et Yangambi km5 (32%). Cette étude démontre le potentiel de production du matériel de plantation de la banana par l’approche in vitro. La capacité de proliferation in vitro et en particulier l’efficacité de produire de pousses recouvrables de différents cultivars pourrait être amélioré en variant les conditions de cultrure Durant les cycles sous culturales. Mots Clés: Banane EA-AAA, In vitro, micropropagation, Musa spp. pousses de bourgeons INTRODUCTION Production of banana (Musa spp.) is limited by scarcity of quality planting materials besides pests, diseases, soil nutrient deficiencies and drought (Baiyeri and Aba, 2005). Conventionally, bananas are propagated by means of corms and suckers which are slow to multiply, bulky and often contaminated with pests and disease pathogens. Bananas with AAA genome composition, including the East African highland bananas (EA-AAA bananas), particularly have low natural proliferation ability leading to inadequate production of suckers to meet farmers’ demand (Baiyeri and Aba, 2005). The increased prevalence of devastating banana bacterial wilt caused by Xanthomonas campestris pv. musacearum, especially in Uganda (Tushemereirwe et al., 2004) has further elevated the demand for quality planting materials. Alternative means for production of banana planting materials are available including macropropagation and in vitro micropropagation (tissue culture). Macropropagation appears to be simpler to conduct by farmers. However, it can propagate infected plants since low cost technologies have not been developed to enable farmers detect disease pathogens from the generated plantlets. Micropropagation has a number of advantages over all other banana propagation methods to both banana breeders and growers. It enables more rapid multiplication of disease-free planting material in a smaller amount of space and has potential to yield several plantlets from a single isolated plant tissue (Vuylsteke, 2001). Micropropagated banana plants also establish more quickly, grow more vigorously, have shorter and more uniform production period, and produce higher yields than propagules produced through other means (Vuylsteke, 2001). In fact, banana plants raised through tissue culture produce more suckers in the field than those propagated conventionally which in another way helps to overcome the problem of shortage of banana planting material (Aish et al., 2003). Micropropagation of banana, especially through shoot tip culture has of recent become a routine technology in many banana growing areas of the world including East Africa. A couple of commercial tissue culture laboratories have been established and the farmer adoption rate of tissue culture plants in the region has been rapidly increasing (AATF, 2003). Consequently, enormous research efforts have targeted enhancing the existing banana micropropagation protocols and adapting them to different EA-AAA banana cultivars. However, there has been scanty information on proliferation potential of bananas in vitro. Under natural field conditions, EA-AAA bananas differ greatly in suckering ability (Karamura and Pickersgill, 1999). Hitherto, there has been no empirical data to compare in vitro proliferation of banana cultivars that naturally have low suckering ability vis-a-vis those with high field suckering ability. Information on in vitro proliferation of EA-AAA banana cultivars would enhance development of micropropagation protocols for their multiplication and aid producers of tissue culture plants in planning, especially in deciding which cultivars would be economic to multiply. The objective of this study was to determine the proliferation potential and the number of recoverable or weanable plantlets that can be generated from a single isolated shoot tip of East African highland banana cultivars with different suckering ability in the field. MATERIALS AND METHODS Banana materials used. Seven EA-AAA banana cultivars (Mpologoma, Musakala, Mbwazirume, Nfuuka, Nakinyika, Nakabululu and Kabula) from different clone sets as described by Karamura and Pickersgill (1999) were studied. Yangambi Km5, an AAA banana, with origin from West Africa (Pillay et al., 2006), and Sukali Ndizi, an exotic banana with genome AAB (Pillay et al., 2003) were included for comparison. Determining in vitro proliferation ability. Shoottip explants of banana cultivars named above were obtained from young suckers of 40-100 cm height, from the banana mother garden at Makerere University Agricultural Research Institute, Kabanyolo (MUARIK). They were trimmed to about 2-3 cm for both diameter and height, and subjected to two processes of sterilisation. They were immersed for 20 minutes in 15 % v/v sodium hypochloride (Jik) to which Tween 20 was added at 0.2 % v/v, and then in 75 % ethanol for 10 minutes before rinsing three times with distilled water. After sterilisation, the shoot-tip explants were reduced to uniform size of 2 cm diameter and height, and carefully bisected longitudinally. The two halves of each shoot tip were initiated in a culture bottle containing 30 ml of the Murashige and Skoog (MS), 1962 medium, modified for banana multiplication as described by Talengera et al. (1994). Fifty shoot-tip explants of each cultivar were initiated and placed in a growth room maintained at 26 ± 2°C, in a light cycle of 16 hr with a photosynthetic photon flux (PPF) of about 60 µmol m-2 s -1 (Strosse et al., 2003). Initially, the cultures were transferred to a fresh multiplication medium, each week for two weeks. After the dead or necrotic parts were cut off at transfer 1 (T1), each explant was split again into two parts; while at transfer 2 (T2) any isolatable shoot meristem was cut and cultured separately. Following T2, sub-culturing was done every 4 weeks for 4 cycles (S1-S4) before transferring the cultures to rooting medium for 4 weeks, and subsequently, weaning them into a pre-steam sterilised soil substrate. At each cycle, proliferating shoots and buds of twenty cultures picked randomly were counted and recorded before a random shoot from each bottle was transferred to the next subculture cycle. Other growth features such as shoot morphology and clustering were also observed and recorded. The number of differentiated shoots and buds were noted, and the total computed as the total shoot proliferation (TSP) potential per cycle. As the shoots multiplied and increased in number with culture cycle, a manageable proportion of twenty to thirty shoots was picked randomly and serially sub-cultured to determine the number of plantlets that could be generated from a shoot tip in a given time period. The number of recoverable or isolatable shoots from an explant, at end of a cycle, was calculated as the ratio of number of plantlets produced at the end of each cycle (P) to the number of shoots subcultured at each cycle (S) to produce P plantlets multiplied by the cumulative number of shoots cultured at that cycle. Derivation of predictive model for recoverable shoots. Based on the non-linear recoverable shoot growth pattern, the data on recovered shoots were transformed into logarithmic scale to create new variables: y = lnY, where: Y is the number of recovered shoots and y is the transformed data. A least squares method was then used to fit the transformed data or new variables to a model using GenStat 12th Edition (Rayne et al., 2009): yij = [(bk + bj)(xi – 1) + bq(xi – 1)2 + c]. Where: yij = is the regression equation of cultivar j
The number of recoverable shoots produced from one shoot tip after a given time could, thus be predicted by converting the prediction for ‘y’ into ‘Y’ as follows: Yij = eyij Yij = e[(bk + bj)(xi – 1) + bq(xi – 1)² + c or Yij = Exp[(bk + bj)(xi– 1) + bq(xi – 1)2 + c]. Where: Yij = number of recoverable shoots producedfrom one shoot tip explant of cultivar j after a given time in weeks The assumptions for this prediction model are that: recoverable shoots of bananas multiply in a linear function followed by a quadratic function; there is no contamination, the error in the transformed equation is normally distributed with mean 0 and other assumptions for regression hold. Data analysis. The data on differentiated shoots, buds and total shoots were subjected to ANOVA using GenStat 12th Edition (Rayne et al., 2009) and the means separated using least significant differences (LSD) at 5% level of significance. The means of the multiplication rates for total shoots and recovered shoots of the cultivars were computed to determine the proportion of total shoots recoverable or effectiveness to generate plantlets from shoot buds. The linear regression coefficients (the rates of growth in number of recoverable shoots) of the cultivars were also separated using LSD at 5% level of significance. RESULTS There was a wide variation in number of differentiated shoots and buds produced in vitro by the different cultivars, with significantly higher numbers among the non EA-AAA banana cultivars (Table 1). All through the subculture cycles, Yangambi Km5 proliferated profusely and produced significantly (P < 0.05) the highest number of shoots among the studied cultivars, followed by Sukali Ndizi. Sukali Ndizi produced significantly (P < 0.05) higher number of shoots than the EA-AAA banana cultivars in most of the subculture cycles. The cultivars with high number of buds in general showed higher proliferation ability. The proliferation trend of the cultivars showed a linear increase and decrease in differentiated shoots and buds, respectively over the successive subculture cycles. The increase in differentiated shoots accounted for 92% of the sum of squares due to cycles, while the decrease in buds accounted for 89% of the sum of squares due to cycles (ANOVA table not shown). In general, the EA-AAA banana cultivars produced between 3 and 4 total shoots, while Sukali Ndizi and Yangambi Km5 produced 5 and 9 total shoots, respectively, at each subculture cycle. Among the EA-AAA banana cultivars, Kabula had higher total shoots, though they were stunted and resulted into fewer recovered shoots. Differences in number of recoverable shoots among the cultivars were recorded as early as transfer 2 at two weeks after the initiation of the cultures (Table 2). By the 4th subculture cycle, the number of shoots recovered from one starting shoot tip ranged from 57 (for Kabula) to 169 (for Nakinyika) among the EA-AAA banana cultivars. Cv. Mbwazirume performed second to Nakinyika with 154 shoots; while the other EA-AAA banana cultivars produced moderate numbers of shoots. Sukali Ndizi and Yangambi Km5 produced significantly more recoverable shoots (322 and 352, respectively) than the EA-AAA banana cultivars. The proportion of total shoots recovered or the ability of the cultivars to produce recoverable shoots was different (Table 3). Shoot recovery ranged from 36% (for Kabula) to 66% (for Mbwazirume) among the EA-AAA banana cultivars. Besides Kabula, all the EA-AAA banana cultivars had relatively high proportion (> 50%) of the total shoots recovered. Yangambi Km5 which had the highest proliferation rate had lower proportion (35%) of total shoots recovered. In general, there was a strong negative correlation between total shoot buds and % recovered shoots (r = 0.94). The morphology and shoot growth characteristics of the cultivars studied were conspicuously different (data not presented). For instance, Yangambi Km5 and Nakinyika cultures had clustered shoots accompanied with many leafy buds while Sukali Ndizi shoots were giant in stature with moderate number of buds. The cultures of cv. Mbwazirume and other EA-AAA banana cultivars had dominant and pointed shoots. The rates of multiplication of recoverable shoots produced from one shoot tip explant of the cultivars were significantly different as exhibited by the linear regression coefficients (Table 4). The EA-AAA banana cultivars had lower multiplication rate of recoverable shoots as compared to Yangambi Km5 and Sukali Ndizi. Among the EA-AAA banana cultivars, Nakinyika, Mbwazirume and Nfuuka had higher multiplication rates followed by Mpologoma, Nakabululu, and Musakala. Kabula had the lowest multiplication rate of recovered shoots. The regression statistic for recoverable shoots had high coefficients of determination (R2 > 90) and thus were used to derive predictive models for the respective cultivars (Table 4). DISCUSSION The EA-AAA banana cultivars showed lower in vitro proliferation rates compared with the non-EA-AAA banana cultivars. The low in vitro proliferation ability of EA-AAA banana cultivars and lack of significant differences in proliferation was reported earlier (Talengera et al., 1994). Besides, this banana group has been reported to be less responsive to regeneration protocols (Sadik et al., 2007) compared to other genotype groups. But the outstanding proliferation rates of Yangambi Km5 (with an AAA genome) and Sukali Ndizi (with an AAB genome) may be of interest to researchers. Conventionally, cultivars having only A in their genome have been reported to produce fewer shoots and buds compared to those having one or two Bs in their genome (Strosse et al., 2003). It is also reported that the origin of the A genome among the EA-AAA bananas is different from that of other commonly grown banana cultivars with A genome (Shepherd, 1957). This could suggest that the A genome of Musa acuminata Colla in the EA-AAA banana is less responsive to in vitro proliferation using the available culture media formulations and protocols. Though cv. Nakabululu did not perform so well over other cultivars after producing a significantly higher number of buds in subculture 1, high bud proliferation especially during the early subculture cycles influenced subsequent proliferation of the cultures, and consequently their overall proliferation ability (Tables 1 - 2). Buds are precursors for shoot development and since they are not fully differentiated, it was likely that they continued to divide when exposed to a fresh medium during sub culturing. This implies that their decrease in number with subculture cycle indicated an inherent overall decline in proliferation rate with increase in subculture cycle or age of the culture. However, as the mean of shoots was still increasing by the fourth subculture cycle, the process of sub-culturing could be continued to produce numerous plants from a single explant. But abnormal plants, especially somaclonal variants might arise and/ or increase to unacceptable numbers due to onset of mutations. Considerable increase in the frequency of variants has been reported in bananas when maintained in tissue culture for over five months or 5 subculture cycles (Rodigues et al., 1998; El-Dougdoug et al., 2007). The age of the culture and or number of subculture cycles affect genetic stability due to spontaneous mutations which accumulate with time (Duncan, 1997; Leela et al., 2003). There was a wide variation in ability of the cultivars to produce recoverable shoots (Table 1). Interestingly, the cultivars with high proliferation rate displayed less efficiency to produce recoverable shoots. This was associated with the number and morphology or growth characteristics of their shoot cultures. For instance, cv. Kabula produced numerous stunted shoots which were difficult to isolate and subculture; while cv. Mbwazirume had moderate number of dominant and pointed shoots which were easy to separate and subculture. Yangambi Km5 produced numerous tiny differentiated shoots that were difficult to recover and subculture compared to Sukali Ndizi with relatively fewer dominant shoots. Since the explants were of uniform size, the variation in number of shoots among the cultivars was probably due to differences in their response to culture conditions. Differences in cultivar proliferation rate have been attributed to differences in growth regulator uptake, translocation rate to meristematic regions, and metabolic processes among genotypes (Blakesley, 1991; Youmbi et al., 2006). The above observations imply that in vitro proliferation ability and, particularly efficiency to produce recoverable shoots of the different EA-AAA banana cultivars, could be improved by varying the culture conditions during the subsequent subculture cycles. Cultivars with a high proportion of buds to differentiated shoots and or numerous tiny shoots require enhanced shoot elongation and dominance. This could be achieved by reducing the amount of cytokinin and other growth inducing substances in the medium in the subsequent subculture cycles, while adding gibberellins especially GA3 which enhances growth and elongation of dwarfed or stunted plantlets in vitro. In bananas, GA3 induced shoot elongation in both genetically dwarf (Damasco et el., 1996) and normal height genotypes (Sanna et al., 2008). In other crops, Veltcheva and Svetleva (2005) achieved desirable shoot elongation and plant recovery from shoot premordia of common bean by applying smaller amount of BAP and higher amount of GA3 in the medium. Similarly, Divya et al. (2008) achieved shoot bud elongation in cotton on medium fortified with reduced amount of BAP but increased amount of GA3. On the other hand, those cultivars with a low proportion of buds to differentiated shoots require enhanced bud proliferation, which could be achieved by increasing the amount of cytokinin and adding other growth inducing substances such as abscisic acid in the subsequent subculture cycles. Application of more active cytokinins with carry-over effect such as Thidiazuron (TDZ) that can sustain bud proliferation in bananas for some time (Makara et al., 2010), might be necessary for those cultivars with very low bud proliferation ability. Cytokinins produce delayed response in undifferentiated tissue (Mineo, 1990) which favours formation of shoot buds. In general, the study suggested that shoot recovery in EA-AAA bananas does not depend on cultivar’s proliferation ability in vitro. Thus, a model for estimating number of plants that could be produced from a shoot-tip explant after a given time, would be useful to those involved in micropropagation of EA-AAA banana cultivars. This study derived a regression model that could be used to predict the number of recoverable plantlets from one shoot-tip explant of EA-AAA banana cultivars after a given time period. Using the predictive model (Table 4), the number of shoots that could be recovered from a shoot-tip explant of cv. Nakinyika after 18 weeks is Y =e[0.289(x-1) – 0.00386(x-1)² +1.39] = 178. The number recorded from the experiment was 169. For cv.Mwazirume, Y = e[0.276(x-1) – 0.00386(x-1)² +1.39] = 142. The one obtained from the experiment was 154. The difference was attributed to changes in shoot growth characteristics of each cultivar. Regression models have been used to predict plant multiplication rates (Mendes et al., 1999) and, thus, the model could be used to give a fair estimate. But the underlining principle, while developing the prediction model, was that bananas may be safely sub-cultured up to the 46th subculture cycle corresponding to 4.5 – 6.5 months. Thus, care must be taken when predicting beyond this period due to possible changes in proliferation or growth pattern depending on cultivar. CONCLUSION In vitro shoot proliferation in East African highland bananas on the widely adopted MS medium modified for banana multiplication, varies from 3-4 new shoots in each subculture cycle and between 57 and 169 recoverable shoots from one starting shoot-tip explant in 18 weeks. The results suggest that in vitro proliferation and recovery of shoots from shoot buds of EA-AAA bananas is cultivar independent. The cultivars that inherently displayed high shoot bud proliferation record low recovered shoots. Based on shoot culture growth characteristics, the number of recoverable shoots from one shoottip explant in a given time can be predicted using a regression model. Using the predictive model, the number of recoverable shoots from one shoottip explant for instance of farmer preferred cv. Mpologoma at 5.5 months (5 subculture cycles) and 6.5 months (6 subculture cycles) would be 176 and 245, respectively. ACKNOWLEDGEMENT The study was funded by World Bank and the Government of Uganda through the Millennium Science Initiative (MSI) Programme administered by the Uganda National Council for Science and Technology (UNCST). REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12015t3.jpg] [cs12015t1.jpg] [cs12015t4.jpg] [cs12015t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}