 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
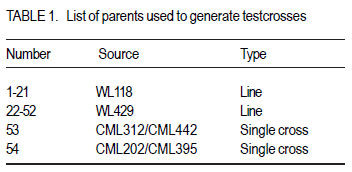
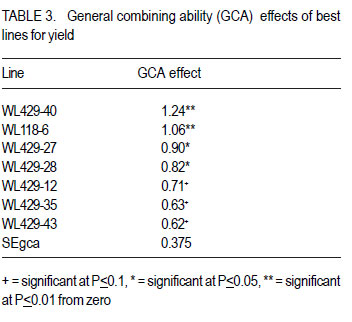
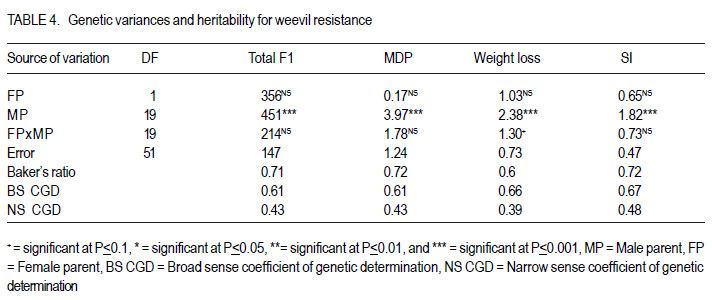
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 99-104 Determination of the heterotic groups of maize inbred lines and the inheritance of their resistance to the maize weevil M. Gafishi Kanyamasoro, J. Karungi1, G. Asea2 and P. Gibson1 Rwanda Agriculture Board, P. O. Box 5016, Kigali, Rwanda Code Number: cs12017 ABSTRACT Maize weevil (Sitophilus zeamais Motschulsky) is a major maize (Zea mays L) storage insect pest in the tropics. Fifty-two inbred lines developed for weevil resistance were crossed to two testers, A and B, to determine their heterotic groups and inheritance of resistance to maize weevil. For 10 testcrosses selected for performance by tester, the correspondent testcrosses onto the opposite tester and all the parents involved in those crosses, were included in the combining study. Of 52 inbred lines, 7 had significantly positive general combining ability (GCA) for yield, with a maximum GCA effect of 1.24t.ha-1 . Based on the specific combining ability (SCA) effect, 23 inbred lines were assigned to heterotic Group A, 24 to Group B, and 5 to both A and B. The combining ability test for weevil resistance revealed that only one inbred, WL118-9, was classified as moderately resistant with a susceptibility index (SI=5.4), two lines were moderately susceptible; and the remaining inbreds were susceptible to highly susceptible to the maize weevil. Additive and non-additive gene action were important for yield, but for weevil resistance, additive gene action was more important. Weevil resistance exists and is moderately heritable. Key Words: Combining ability, gene action, Sitophilus zeamais, susceptibility index, Zea mays RÉSUMÉ Le charançon du maïs (Sitophilus zeamais Motschulsky) est un plus grand ravageur du maïs (Zea mays L) dans les tropiques. Cinquante-deux lignées développées pour la résistance au charançon du maïs étaient croisées aux testeurs A et B afin de déterminer leurs groupes hétérotiques ainsi que la transmission de la résistance au charançon du maïs. Pour dix lignées sélectionnées pour test de performance par le testeur, les croisements correspondants au testeur opposé ainsi que tous les parents impliqués dans le croisement étaient inclus dans l’ étude d’aptitude. Parmi les 52 lignées, 7 avaient significativement montré une positive Aptitude Générale à la combinaison (GCA) pour le rendement, avec un effet maximum GCA de 1.24 t ha-1 . Basé sur l’ Aptitude Spécifique à la Combinaison (SCA), 23 lignées ont été attribuées au groupe hétérotique A, 24 au groupe B et 5 aux deux groupes hétérotiques A et B. Le test d’aptitude à la résistance au charançon a révélée q’une seule lignée, WL118-9 était classifiée comme modérément résistante avec un index de susceptibilité de 5.4, deux lignées étaient modérément susceptibles tandis que les lignées restantes étaient soit susceptibles ou hautement susceptibles au charançon du maïs. L’action additive et non additive des gènes étaient très importante. Les gènes de résistance au charançon du maïs existent et sont modérément transmissibles à la descendance. Mots Clés: Aptitude à la combinaison, action des gènes, Sitophilus zeamais, index de susceptibilité, Zea mays INTRODUCTION Maize (Zea mays) plays an important role in the diet of millions of African people (Fandohan et al., 2003). In developed countries, it is changing from food to a high-value economic crop. Maize ranks first in terms of production among cereals, ahead of wheat and paddy rice. The crop however, is prone to various production constraints including storage pests. Among the key pests in storage, the maize weevil Sitophilus zeamais is one of the major maize constraints affecting production especially for susceptible genotypes. This insect pest is classified as primary category of grain insects due to its ability to destroy a whole sound grain. Both adults and larvae are internal grain feeders and cause huge losses to stored maize. A loss of up to 80% has been reported by Dlhiwayo and Pixley (2003) on untreated maize stored in traditional structures in most of tropical countries due to the maize weevil. This insect pest not only destroys the grain but also contaminates it with insect byproducts that significantly affect the grain quality and render it unsuitable for human consumption. In addition to direct destruction of the maize grain, wounded grains are favorable to fungal infestation and subsequent contamination of mycotoxins associated with cancer in humans (Presello, 2006). Weevilled grains are also widely known to fetch low prices in the market. Although some measures to control insect infestation exist, these are not cost effective to resource-poor farmers due to their costs and risk to unsophisticated users and environment. The indiscriminate use of chemicals was reported by Gu et al. (2008) to lead not only to development of resistance in invertebrate pests, but also to give raise to secondary insect species due to the destruction of their natural enemies in the ecosystem. Host plant resistance can help to suppress the insect population to below grain damaging levels, and is therefore, a very important integrated pest management strategy suitable for poor-resource farmers. Genotypes resistant to maize weevil are still scarce (Derera et al., 2010). Breeding has been for long focused on improvement of performance and tolerance to field biotic and abiotic stresses ignoring traits that improve grain storage. Weevil resistance exists though its inheritance is low (Kim and Kossou, 2003). Bergvinson (2000) stated that the low heritability of weevil resistance implies slow progress in moving the trait into elite genotypes. The relative importance of additive and nonadditive gene action is a prerequisite in determining how building the trait into elite genotyes can be achieved. A number of research works on weevil resistance suggested that additive and non-additive gene action is important for this trait (Derera et al., 2001; García-Lara et al. 2009). Heterosis in maize is well documented and has made maize improvement a success through hybrid production. Good inbred lines that combine both yield and weevil resistance will significantly contribute to suppressing the weevil population in the storage facilities, reduce dependence on pesticides and improve income, especially for resource-poor farmers. Understanding the inheritance of resistance to insects and determining the heterotic groups of inbreds should ease the selection of lines to be used for the development of superior hybrids with good resistance to maize weevils. MATERIALS AND METHODS Description of the research area. The research was conducted in two locations, Namulonge and Masaka. Namulonge is located within the bimodal rainfall region. It lies at 00 32'’ N of the Equator and 320 37'’ E and located at 27 km North of Kampala the capital city of Uganda, at an elevation of 1150 masl. It has a tropical wet and mild dry climate (800 to 1,100mm annual precipitation) with slightly humid conditions (average 65%) and a mean temperature of 270C. Masaka’s annual rainfall ranges from 1000-1500 mm with average temperature of 15oC to 27oC and >80% relative humidity (Lipps et al., 1997). Nursery and seed production. Seeds of 104 testcrosses were developed from a Line x Tester mating design of 52 inbred lines and 2 single cross hybrid testers, A (CML312 x CML442) and B (CML202 x CML395) at the National Crops Resources Research Institute (NaCRRI), Namulonge 2009A (Table 1). The planting distances were 0.75m between rows and 0.30m between hills on a two-5m row plot for each inbred line. The testers were planted at 27 rows each at the same density as the inbred lines. Shoot and tassel bagging were carried out before silk emergence and pollen shed to avoid contamination followed by hand pollination. The inbred lines were the male parents and testers the female parents in those crosses. Evaluation for field performance. The 104 testcrosses were planted on a two-5m row plot each at Namulonge and Masaka, 2009B cropping season under rain-fed condition. The experiment was in 9 x 12 alpha-lattice design replicated twice. Di-Ammonium phosphate and Urea were the mineral fertilisers applied at a rate of 100 kg ha-1 for both fertiliser types. Early and high-yielding testcrosses that are disease resistant having a reduced height and ear placement with good husk cover were selected for maize weevil resistance evaluation. For 10 testcrosses selected per tester, the corresponding testcross onto the opposite tester was also included to allow analysis of combining ability for weevil resistance. The parents involved in those crosses were also evaluated and all of other inbreds as long as they yielded enough seed for evaluation. Evaluation for maize weevil resistance. Fifty newly hatched maize weevils collected from a mass rearing unit of the entomology laboratory were used to infest 100 grams of Longe 5 (an open pollinated QPM variety) in glass jars with perforated lids sealed with wire mesh for insect rearing. A sample of 50 grams of untreated sound grains obtained from the selected germplasms were wrapped in polythene bags and frozen at 20oC for 2 weeks to kill any insect /egg that could have randomly attacked the grains. After this period of time, the samples were transferred to evaluation jars and left for 3weeks to achieve uniform temperature and moisture content before infestation. The lids of the 250 ml evaluation jars were sealed with wire mesh and perforated with 4 large holes for maximum aeration and total insect containment. A total of 32 unsexed insects were used to infest a 50 gram grain sample (Derera et al., 2001; Dhliwayo et al., 2003; Dhliwayo et al., 2005; Derera et al., 2010) as recommended for a rapid screening of genotypes for maize weevil resistance. The samples were given a 10-days oviposition period after which all adult insects were carefully removed, dead and living counted. The samples were then left for incubation at room temperature. The jars were arranged in a 9x10 alpha lattice with three replications. After 32 days of incubation, the visible F1 insect progeny were counted and removed from the jars every other day until no more insect emerged. A Thermo-Hygro clock M288CTH hygrometer type was used to monitor the humidity and temperature status during the experiment period. The total F1insect emergence, the loss of weight and Dobie’s susceptibility index were the insect resistance parameters used in this study. The cumulative weevil number was recorded for the duration of the total F1 insect generation emergence of each genotype. At the end of experiment, the samples were vigorously sieved to remove the flour resulting from insect feeding and tunneling for estimation of the grain weight loss that was obtained by subtracting the final grain weight from the initial weight. Time to emergence of 50% of F1 also called median development period (MDP) was estimated as the number of days from day 5 of oviposition to 50% emergence of progeny. Dobie’s susceptibility index (SI) was calculated using the following equation: SI= (Log F1 x 100)/MDP. Analysis of variance for heterosis was performed using linear mixed models from GENSTAT 12th Edition to determine the variance of the testcrosses, ANOVA for combining ability to ascertain the contribution of parents and their interaction to the hybrid performance. ALLPAIRWISE procedure of GENSTAT was used for mean separation and assessment of performance in yield. General combining ability effect was estimated for inbred lines as mean of all crosses involving the male parent minus grand mean. The significance of GCA effect was obtained computing standard error for male parent and tested against t-test using the GxE degrees of freedom. Evaluation of testcrosses for weevil resistance used only seed from Namulonge site. The relative importance of general combining ability (GCA) and specific combining ability (SCA) was determined using Baker’s ratio (2GCA/2GCA+SCA) (Baker, 1978). The broad-sense and narrow-sense coefficient of genetic determination were calculated using the formulas 2GCA+SCA/2GCA+SCA+Error and 2GCA/2GCA+SCA +Error, respectively RESULTS AND DISCUSSION There were significant differences for yield averaged across environments. The GxE was highly significant for grain yield implying that the testcrosses performed differently in different environments. Derera et al. (2008) found similar results. In this study, TA/WL429-40, TA/WL42939, TA/WL429-27, TA/WL429-35 were among the testcrosses with good performance in both locations. The genetic variances across sites revealed a highly significant contribution of both female and male parents to increased yield and a significant role of the interaction (Table 2). Heritability for yield was low (Table 2). A Baker’s ratio of 0.56 for grain yield, suggests that additive genes were slightly more important than non-additive for the control of yield. Only 7 inbred lines of 24 with positive GCA values for yield were significantly greater than zero with 2 exceeding a GCA of 1.0 t ha-1. These lines were the best combiners for yield (Table 3). The remaining male parents exhibited either nonsignificant positive or negative GCA effects for grain yield, meaning average to low contribution to yield. Some of the best combiners for yield were however associated with unfavorable effects for important traits. Most of the lines except one (WL429-28) had some detrimental effects on traits other than yield indicating the difficulty in obtain inbreds with desirable expression of all the traits of importance. A desirable cross is one in which the parents are genetically divergent and produce progeny with good performance, and which exhibits a high specific combining ability (Xingming et al., 2001; Legesse et al., 2009). The two testers A and B successfully discriminated among the 52 inbred lines of unknown heterotic pattern. An inbred was assigned to group A when its cross with A showed a large negative SCA value otherwise it was assigned to group B. Based on this criterion, 23 inbred lines were assigned to the heterotic group A, 24 assigned to heterotic group B, and 5 inbred lines that showed similar performance when crossed with tester A or tester B were assigned to both heterotic groups, A and B. Analysis for weevil resistance revealed that there were highly significant differences among entries for all resistance parameters, confirming the presence of resistance mechanisms at different levels among the genotypes. High total F1 progeny emergence, high grain weight loss and high SI were quite strongly related (P<0.001, r2 > 0.75). Correlations among the 4 resistance parameters were highly significant and strong association was especially with SI. Derera et al. (2010) found strong positive correlation between the number of progeny and kernel damage, and between the number of progeny per adult and grain weight loss per adult. Dobie index classifies genotypes into 5 resistance categories: SI<4 for Resistant, SI = 4.1 to 6 for moderately resistant, SI = 6.1 to 8 for moderately susceptible, SI = 8.1 to 10 for susceptible and SI>10.1 for highly susceptible genotypes. Based on this classification, only 1 line WL118-9 with SI of 5.4 was classified as moderately resistant (MR) and best combiner for weevil resistance. The lines WL118-14 and WL429-27 (SI=7.3) were moderately susceptible to maize weevil. The remaining inbred lines were susceptible to highly susceptible. Further correlation evaluation was between grain texture and the resistance parameters used in this study. Flint texture is reported to dissuade weevils from feeding and laying eggs. On the contrary, dent texture indicates a soft kernel endosperm easy for weevils to damage (Sadivan, 2002; Kim and Kossou, 2003; Hossain et al., 2007). A dent type of grain texture (high score) was weakly correlated with a low MDP (P<0.1). Low time of 50% F1 emergence indicates little time for weevils to destroy the grains and therefore the susceptibility of the genotype. Weak association implies that grain texture alone cannot predict weevil resistance. The other measures of resistance did not correlate with grain texture. This is in agreement with Demissie et al. (2008) who stated that flint grains are more resistant than dent, but texture alone is not a sufficient indicator of weevil resistance. Though the experiment was conducted at room temperature, we were able to identify lines that carry weevil resistance. The temperature ranged from 24oC to 26oC and humidity 65% to 75% during the experiment, slightly low compared to optimum temperature and humidity of 28±2oC and 70±5% often used for the insect in various research works. The WL118-9 was the best general combiner for weevil resistance of all the inbred parents used in this study, but it had only a very small positive GCA effect for yield (0.10 t ha-1). The WL429-27 had a significant GCA effect for yield and has fairly contributed to weevil resistance. These lines can be used to improve yield and weevil resistance. The lines harbored and transmitted weevil resistance to progeny. Male parents contributed more to weevil resistance than female parents and their interaction (Table 4). There was no highly resistant inbred line among the studied lines. Additive and non additive gene action were important for yield, but additive gene action was more important than non additive for weevil resistance. (Baker’s ratio>60 for all resistance parameters) (Table 4). These data reveal that weevil resistance exists in the tested maize genotypes and is moderately heritable. ACKNOWLEDGEMENT This study was funded by Strengthening Capacity for Agricultural Research and Development in Africa (SCARDA) through Regional Universities Forum for Capacity Building in Agriculture (RUFORUM). The studies were implemented by various institutions and the authors are grateful to Makerere University for the training, the National Crops Resources Research Institute (NaCRRI) for providing required facilities and technicians and Rwanda Agriculture Board (RAB) for all the support. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12017t2.jpg] [cs12017t3.jpg] [cs12017t4.jpg] [cs12017t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}