 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 105-112 Genetic analysis of resistance to rice bacterial blight in Uganda I. Habarurema, G. Asea1, J. Lamo1, P. Gibson, R. Edema, Y. Séré 2 and R.O. Onasanya3 College of Agricultural and Environmental Sciences, Makerere University, P. O. Box 7062, Kampala, Uganda
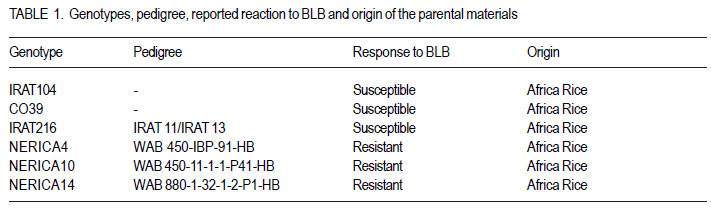
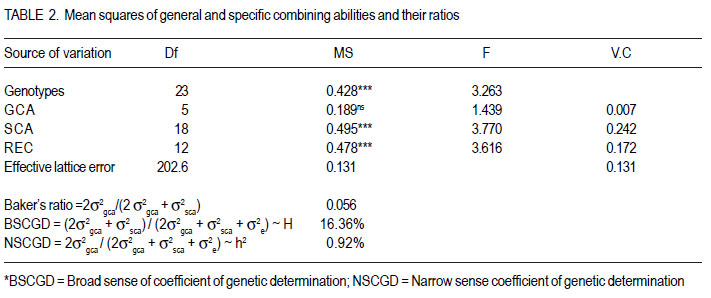
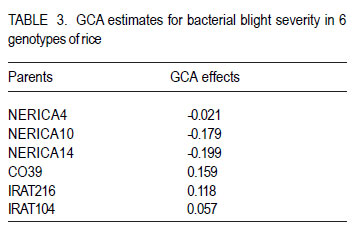
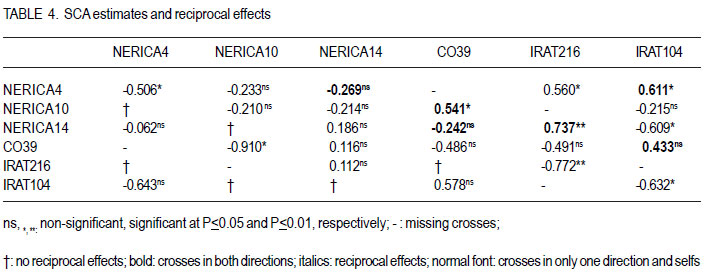
Code Number: cs12018 ABSTRACT Rice bacterial blight (Xanthomonas oryzae pv.oryzae) is a major constraint to rice (Oryza sativa L.) production in Uganda and as part of strategies to develop resistant cultivars, it is important to evaluate resistance of commonly used cultivars. A full-diallel mating design involving three resistant and three susceptible rice cultivars was used to produce F1 and F2 progenies in a screen-house at the National Crop Resources Research Institute (NaCRRI), Namulonge in Uganda. The parents and F2 populations were challenged with the Xanthomonas oryzae pv.oryzae isolate (UX00) and lesion lengths were scored 21 days after inoculation (DAI). Griffing’s combining ability analysis showed significant specific combining ability (SCA) and non-significant general combining ability (GCA) effects, indicating the preponderance of non-additive gene effects in controlling the resistance to bacterial leaf blight (BLB) in rice. Rice genotypes, NERICA14, NERICA10 and NERICA4 had desirable GCA estimates, and were, therefore, the best general combiners. Crosses CO39 x NERICA10 and NERICA14 x IRAT104 had favorable SCA values. These hybrids are thus, promising in developing the BLB resistant progenies. Significant reciprocal effects indicate the importance of maternal contribution in controlling the BLB virulence. For this, resistant lines can be used as female parents for fear of affecting transfer of resistance to the progenies, and the hybrids and their reciprocals would be handled separately. Low estimates of narrow sense coefficient of genetic determination (NSCGD) (0.9%) and medium broad sense coefficient of genetic determination (BSCGD) estimates (16.4%) highlight the influence of non-additive gene action in controlling the resistance to BLB, confirming an effective selection of superior genotypes at advanced generations when the maximum homozygosity is fixed. Key Words: Gene action, Oryza sativa, Xanthomonas oryzae RÉSUMÉ Le flétrissement bactérien des feuilles du riz (Xanthomonas oryzae) constitue une contrainte majeur à la production du riz (Oryza sativa L.) en Uganda. L’évaluation de la résistance des cultivars communément utilisés s’avère nécessaire comme une des strategies de développement des cultivars résistants. Des cultivars de riz (Oryza sativa L.) dont trois résistants et trois susceptibles étaient croisés en dispositif diallèle complet pour produire des générations F1 et F2 dans une serre de l’ Institut National de Recherche sur les Resources Végétales (NaCRRI) à Namulonge en Ouganda. Les parents et les populations F2 étaient soumis à l’isolat UX00 du Xanthomonas oryzae pv.oryzae et les longueurs de la lésion étaient évaluées 21 jours après l’inoculation. L’analyse de l’aptitude à la combinaison par la méthode de Griffing ont révélé des effets significatifs de l’Aptitude Spécifique à la Combinaison (ASC) et non significatifs de l’Aptitude Générale à la Combinaison (AGC), indiquant la prépondérance des effets génétiques non additifs dans le contrôle de la résistance au flétrissement bactérien des feuilles (FBF). Des génotypes NERICA14, NERICA10 et NERICA4 avaient des valeurs souhaitables d’AGC, et étaient ainsi des meilleurs combinants pour la résistance au FBF. Des hybrides CO39 x NERICA10 et NERICA14 x IRAT104 avaient des valeurs favorables d’ASC. Ils sont donc promettant dans le développement des descendants résistants au FBF. Des effets réciproques significatifs indiquent l’importance de la contribution maternelle dans le contrôle de la virulence du FBF. Pour ce faire, des lignées résistantes peuvent être utilisées comme parents femelles de peur d’affecter le transfert de la résistance aux descendants, et les hybrids ainsi que leur réciproques pourraient être manipulés séparément. De faibles valeurs (0.9%) du coefficient génétique de détermination au sens restreint et valeurs moyennes (16.4%) du coefficient génétique de détermination au sens large accentuent l’influence de l’action génétique non additive dans le contrôle de la résistance au FBF, ce qui confirme une sélection effective des meilleurs génotypes aux générations avancées quand le maximum d’homozygotie est fixé. Mots Clés: Action génétique, Oryza sativa, Xanthomonas oryzae INTRODUCTION Rice bacterial leaf blight (BLB) caused by Xanthomonas oryzae pv.oryzae, is one of the most devastating diseases of irrigated and rainfed lowland rice (Oryza sativa L.) worldwide (Mew, 1987; Chu et al., 2006), and contributes to substantial yield losses of up to 50% (Adhikari et al., 1995; Gnanamanickam et al., 1999). Several approaches that have been used to combat bacterial leaf blight include cultural practices, biological and chemical controls. However, the cost-effective way to control this disease among resource-poor farmers is the use of resistant rice cultivars. Unfortunately, the bacterial leaf blight pathogen exists with a very high degree of race-cultivar specificity (Niño-Liu et al., 2006). Over 30 races of X. oryzae pv.oryzae have been reported in several countries (Adhikari et al., 1999b; Shanti et al., 2001) and new races of the pathogen continue to evolve and overcome the deployed resistance genes (Mew, 1987; Mew et al., 1992). Most of these races differ between regions, sites and even fields within a site (Nelson et al., 1996). This implies that it is important for breeding for resistance to BLB to be location specific using the prevailing races in that particular geographical location. To date, over 30 resistant (R) genes against the bacterial blight pathogen have been identified in rice (Liu et al., 2007) and designated in a series from Xa1 to Xa31 (Wang et al., 2009; Lore et al., 2011). A number of these genes are dominant, for example, Xa1, Xa2, Xa3, Xa4, Xa6, Xa7, Xa10, Xa11, Xa12, Xa14, Xa16, Xa17, Xa18, Xa21, Xa22, Xa23, Xa25, Xa27, Xa29(t) and Xa31(t) (Niño-Liu et al., 2006; Wang et al., 2009; Lore et al., 2011); whereas genes, xa5, xa8, xa9, xa13, xa15, xa19, xa20, xa24, xa26b, xa28 and xa32(t) are considered recessive genes (Kameswara et al., 2002; Iyer-Pascuzzi and McCouch, 2007; Shanti et al., 2010). All these genes have become the main sources for the genetic improvement of rice for resistance to X.oryzae pv.oryzae worldwide. Unfortunately, none of the isolate from Uganda has been used in studying the genetics of resistance to BLB in the cultivars presently cultivated in the country. Furthermore, knowledge of combining ability is essential for selection of suitable parents for hybridisation and identification of promising hybrids in breeding programme. This study sought to establish the resistance to the BLB isolate from Uganda using the upland rice varieties NERICA4, NERICA10 and NERICA14, and other local lowland cultivars available in Uganda. MATERIALS AND METHODS The study was conducted at National Crops Resources Research Institute (NaCRRI)-Namulonge, located at approximately 27 km North Kampala City along latitude 0°32’N and longitude 32°53’E, from August 2009 up to November 2010. Choice of parents. Six parental genotypes used were obtained from AfricaRice and were chosen based on the reaction to Namulonge BLB isolate. They comprised three resistant and three susceptible lines. The resistant lines were upland NERICAs, namely, NERICA 4, 10 and 14, all already released to the Ugandan farmers and seed growers; while the susceptible lines included IRAT lines 104 and 216, and CO39 that were elite lines still under evaluation (Table 1). Hybridisation and progenies handling. Seeds were sawn in wooden boxes in hills with 15 cm x 20- cm intra- and inter-row spacing at the rate of three to five seeds per hill, but later thinned to only two seedlings. Planting of the parents was done under screenhouse conditions and was staggered at two to three week intervals in three sets to enable synchronisation of the flowering dates, and subsequently allow a successful crossing (Sarkarung, 1991). At flowering stage, all the parental lines were crossed in the screenhouse using the full-diallel mating design in which crosses were made in all possible combinations, including reciprocals (Griffing, 1956a, b). The resultant F1 seeds were harvested, sun-dried, planted and selfed to generate F2 segregating populations. Seeds of F2 progenies and parents were dried and planted in plastic pots at the rate of five seeds per pot before thinning to three plants one month after germination. Cultural practices like watering, weeding, fertiliser and pesticide application were followed as recommended (Adhikari et al., 1995; 1999a, b; Mannan et al., 2009). Experimental design and progeny assessment. A total of 24 genotypes were randomly planted in alpha-lattice design, with 10 replications. The experimental unit consisted of one pot of three plants. The 24 genotypes used in the study included 18 F2 populations, namely, NERICA4 x NERICA10, NERICA10 x NERICA14, NERICA4 x NERICA14, NERICA10 x CO39, NERICA14 x CO39, CO39 x IRAT216, NERICA4 x IRAT216, NERICA14 x IRAT216, IRAT104 x NERICA10, NERICA4 x IRAT104, NERICA14 x IRAT104, CO39 x IRAT104, CO39 x NERICA10, CO39 x NERICA14, IRAT216 x NERICA14, IRAT104 x NERICA4, IRAT104 x CO39, NERICA14 x NERICA4 and six parents, namely, IRAT216, NERICA4, NERICA10, IRAT104, NERICA14 and CO39. At maximum tillering to booting stage (60 days after planting), three fully expanded leaves per plant were clip-inoculated (Kauffman et al., 1973) with a bacterial suspension containing 108 cells ml-1 using a sterilised pair of scissors. Three plants were inoculated per pot, thus, making a total of nine leaves inoculated per pot. The inoculum used was from a bacterial culture of UX00 as a representative isolate, grown on PSA medium and incubated at 28 °C for 3 days (Lee et al., 2003; Mannan et al., 2009). Inoculated plants were grown under optimum day temperature of 23.4 to 30.6 °C and relative humidity of 54.0 to 68.7%, both measured over 3-week-infection period using a thermo-hygroclock M288CTH. The plants were sufficiently and regularly watered to avoid any interference of drought stress with bacterial blight symptoms. The length of lesions caused by BLB was measured on each inoculated leaf 21 days after inoculation (Gonzalez et al., 2007). The mean lesion length for the plant was the average of the lesion lengths from the three inoculated leaves and was used to assess resistance or susceptibility of individual plants. Plants with score lower than 3 cm were classified resistant, whereas those with higher score were classified susceptible (Deng et al., 2006). Combining ability analysis. Data analysis was carried out using GenStat 12th Edition (GenStat, 2010). The variance was partitioned into the general combining ability (GCA), the specific combining ability (SCA) and the reciprocal effects (REC), according to Griffing’s (1956b) model I, method I modified for missing crosses (P. Gibson, Pers. com, July 2011, Makerere University). The following statistical model was used Xijk = u + gi + gj + sij + rij + 1/bcÓÓeijkl where, Xijk is the observation value for a cross between the parents ith and jth; u is the population mean; gi and gj are the GCA effects for the ith and jth parents, respectively; sij is the SCA effect for the hybrid between the ith and jth parents such that sij = sji ; rij is the reciprocal effect involving the reciprocal crosses between the ith and jth parents such that rij = - rji , and eijkl is the environmental effect associated with the ijklth individual observation. Significant differences among GCA effects and SCA effects were tested using F-values. The parents were considered as a fixed set of lines deliberately selected based on their resistance or susceptibility to the bacterial blight (fixed model). Method I including parents, F1’s and reciprocals was employed because accurate information on reciprocal effects was needed. To adjust the missing crosses, weighted mean of each parent were used to estimate the GCA effect of that parent. Variance components for GCA and SCA were calculated by using σ2gca and σ2sca coefficients appropriate for missing crosses, as suggested by P. Gibson (Pers. com., July 2011, Makerere University). Two-tailed t-tests were used to determine the significance of the GCA and SCA effects. Significant and negative GCA and SCA effects were taken as beneficial, nonsignificant and negative as average and positive as detrimental (Saleem et al., 2010). To explain the parental performance, ratios of mean square components associated with variance of GCA and SCA effects were computed to estimate the relative importance of GCA, as suggested by Baker (1978) with the following formula: 2σ2 gca /2 σ2 gca + σ2 sca such that the closer the ratio is to unity, the greater the predictability of progeny performance based on GCA effects alone. Furthermore, estimates were obtained for the broad sense coefficient of genetic determination (BSCGD), the fixed effect equivalent of broad sense heritability) and narrow sense coefficient of genetic determination (NSCGD), the fixed effect equivalent of narrow sense heritability) for resistance to BLB, as follows: BSCGD = 2ó2gca+ ó2 /2 ó2 + ó2 + ó2 ~ H and and NSCGD = 2ó2 gca /2 ó2 gca + ó2 sca + ó2 e ~ h2 (Jacquard, 1983; Abney et al., 2000). RESULTS AND DISCUSSION The partitioning of the variance into the GCA of the parents, the SCA and reciprocal effects (REC) of the crosses, and their respective ratios are presented in Table 2. Significant mean squares were observed for SCA, but not GCA (Table 2). Additionally, the SCA mean square was about 2.5 times higher than that of the GCA. However, previous studies showed that both GCA and SCA effects were important in governing resistance to BLB (Tabien, 1989), which is the contrary to the present results. Significant SCA effects suggest high contribution of non-additive genetic component in controlling the bacterial blight resistance in the rice genotypes evaluated, whereas the lack of significant GCA effects among genotypes used in this study suggests that additive effects were not expressed for BLB infection (Kenga et al., 2004). Individual GCA estimates are presented in Table 3. The GCA effects were not significant (P< 0.05) for any of the 6 parents. Negative and lower GCA effects are desirable in selecting superior parents for resistance to the BLB because they indicate a larger contribution to the resistance; while positive values imply a contribution towards susceptibility (Kenga et al., 2004; Saleem et al., 2010). The parent, NERICA14, had the lowest negative GCA value (-0.199), followed by NERICA10 (-0.179) and NERICA4 (-0.021); whereas the parent CO39 had the highest positive GCA value (0.159), followed by IRAT216 (0.118) and IRAT104 (0.057). This suggests that the NERICA lines, namely NERICA14, NERICA10 and NERICA4, were identified as the best general combiners for resistance to the BLB and their use in breeding programmes would produce progenies with increased resistance (Kenga et al., 2004); whereas the IRAT lines 104 and 216, and CO39 were the worst general combiners because of their positive GCA values. Negative SCA effects were also desirable for resistance. The highest SCA effect was - 0.91 (P < 0.05) produced in the cross involving CO39 x NERICA10, followed by that produced from a cross involving NERICA14 x IRAT104 (SCA= 0.609; P<0.05), as indicated in Table 4. Hybrids CO39 x NERICA10 and NERICA14 x IRAT104 were therefore identified as the most promising in breeding programmes for resistance to BLB in rice. These hybrids were originated from at least one parent that had a desirable GCA value, namely, NERICA10 or NERICA14. However, the parents with negative GCA effects did not always form good hybrid combinations. For instance, the hybrids NERICA4 x IRAT216, NERICA10 x CO39, NERICA4 x IRAT104 and NERICA14 x IRAT216 involving at least one good combiner (NERICA4, NERICA10 and NERICA14, respectively) were undesirable. Kenga et al. (2004) suggest that the difficulty in predicting the resistance level of the hybrid, on the basis of GCA alone should necessitate testing of specific male-female combinations. The SCA values provide important information about the performance of the hybrid relative to its parents. However, Arunga et al. (2010) found that the SCA effect alone has limited value for parental choice in breeding programmes. They, therefore, suggested that the SCA effects should be used in combination with other parameters, such as hybrid means and the GCA of the respective parents such that a hybrid combination with both high mean and favorable SCA estimates and involving at least one of the parents with high GCA, would tend to increase the concentration of favorable alleles; which is desired by any breeder. Furthermore, it was observed that crosses involving one good combiner and one average or poor combiner showed negative SCA effects. For example, IRAT104 and CO39 had poor GCA values for disease resistance, while their crosses with NERICA14 and NERICA10, respectively, had significant and desirable SCA effects. Hannan et al. (2007) observed that some parents exhibited a similar phenomenon in studies on tomato (Lycopersicum esculentum Mill.). Moreover, highly significant reciprocal effects for the BLB resistance exhibited the importance of maternal contribution of the parents used in hybridisation. Thus, the maternal effects direct the inheritance of resistance to BLB. This suggests that hybrids and their reciprocals should be separately handled and resistant lines should be used as female parents in crosses to generate BLB-resistance in breeding populations. The combining ability ratio, also known as Baker’s ratio, for resistance to the bacterial blight was 5.6% (Table 2). According to Baker (1978), when combining ability ratio approaches unity, GCA alone can predict the performance of the parents. Thus, the GCA scores could not be used to predict the performance of the parents in the present study, because the value of Baker’s ratio is much lower than the theoretical maximum of unity. Low Baker’s ratio highlighted the importance of SCA variance, and hence the importance of dominance and/or epistatic gene effects in increasing resistance to the bacterial blight in rice (Griffing, 1956a; Christie and Shattuck, 1992). This implies that late selection would be done much more based on better hybrid combinations rather than the performance of the parents involved in crossing programmes. In the present study, the values of narrow sense coefficient of genetic determination (NSCGD) and broad sense coefficient of genetic determination (BSCGD) were very low and represented 0.9 and 16.4%, respectively (Table 2). Thus, BSCGD was about 18 times higher than NSCGD. It follows that resistance to the bacterial blight in rice was much more heritable in broad sense (H) than in narrow sense (h2) and that the greater portion of heritable variation is of nonadditive nature. This is in contradiction with the findings of Tabien (1989) that narrow sense heritability was high across races, ranging from 62 to 96%, when four different races from Philippine were tested on eight rice varieties. Therefore, the low Baker’s ratio value and the importance of broad sense heritability relative to narrow sense heritability emphasized the preponderant role of non-additive gene action in controlling the bacterial blight virulence. CONCLUSION Non-additive gene action was preponderant in governing the resistance to the BLB in rice. Genotypes, NERICA14, NERICA10 and NERICA4 are the good general combiners for reducing the BLB severity in rice. Hybrids NERICA14 x IRAT104 and CO39 x NERICA10 are recommended for utilisation in hybrid rice development, because they exhibit favorable SCA estimates and involve the parents NERICA14 and NERICA10, respectively as good combiners. The importance of maternal contribution of the parents used in hybridisation suggests the importance of handling separately the hybrids and their respective reciprocals for selection, and a critical choice of parents for this trait. Low heritability estimates highlight the important influence of non-additive gene action in the expression of the resistance to the BLB in rice, which implies selection of the best lines at later generations to allow fixation of maximum homozygosity. ACKNOWLEDGEMENT The financial support of the Alliance for a Green Revolution in Africa (AGRA) is greatly acknowledged. The National Crops Resources Research Institute (NaCRRI) staff at Namulonge is thanked for hosting the research work and kindly allowing access to various facilities. AfricaRice is acknowledged for providing seeds of different plant materials used in this study. The Regional Universities Forum for Capacity Building in Africa (RUFORUM), Makerere University staff and the Director General of Institut des Sciences Agronomiques du Rwanda (ISAR) provided close collaboration in a bid to strenghten the capacity building in the region. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12018t1.jpg] [cs12018t3.jpg] [cs12018t2.jpg] [cs12018t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}