 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 155-161 Inheritance of resistance to turcicum leaf blight in sorghum M.M. Beshir1, 2, A.M. Ali2 and P. Okori1 1College of Agricultural and Environmental Sciences, Makerere University, P. O. Box 7062, Kampala, Uganda
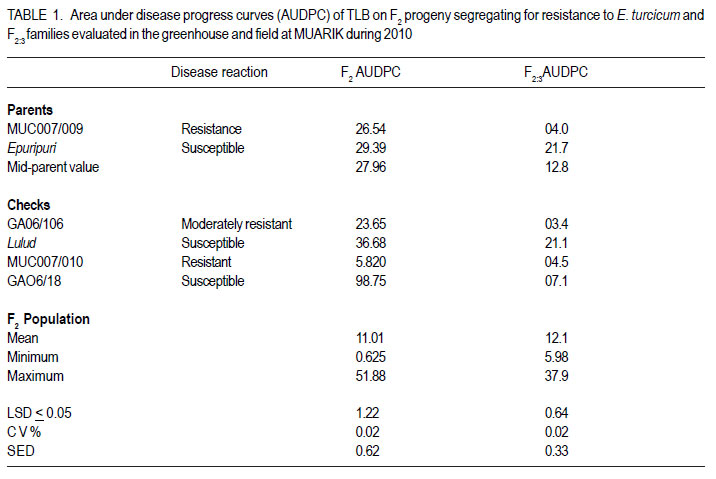
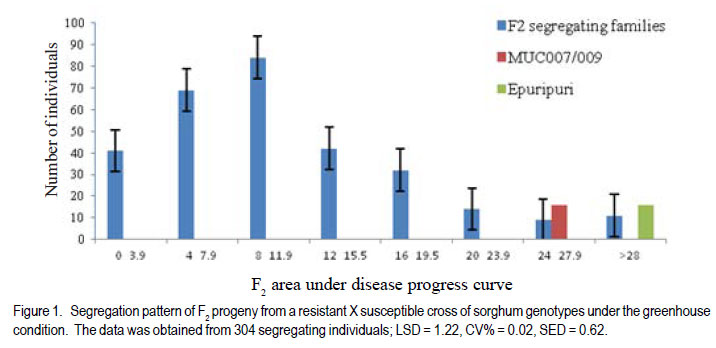
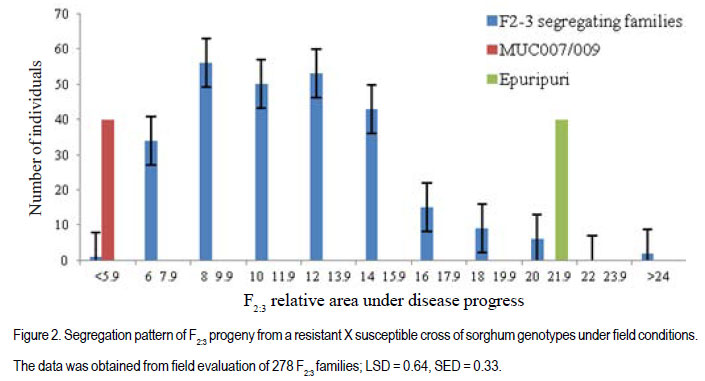
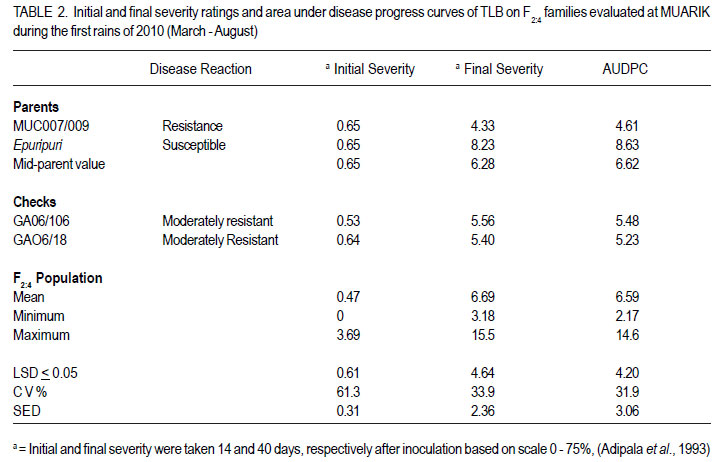
Code Number: cs12022 ABSTRACT Sorghum (Sorghum bicolor) has global socio-economic importance and is as model plant species for many tropical grasses with complex genomes. It is frequently devastated by Turcicum leaf blight, caused by Exserohilum turcicum, leading to considerable grain and fodder yield losses. Developing varieties with resistance against E. turcicum is the most cost-effective way to manage the disease. However, inheritance of resistance to E. turcicum in sorghum is poorly understood. Studies were carried out in Uganda to investigate the mode of inheritance of resistance to E. turcicum in sorghum under greenhouse and field conditions. Segregating families derived from a cross of MUC007/009 (a local resistant accession) and Epuripuri (susceptible, an elite sorghum variety) were used along with the two parents in the study. Evaluations of families also included four checks, namely GAO6/ 106 (moderately resistant), Lulud (susceptible), MUC007/010 (resistant) and GAO6/18 (moderately susceptible). Disease severity of F2 plants in the greenhouse were skewed toward resistance. In the field, the resistant parent had much lower disease severity than the susceptible parent. However, there was no difference between both parents under greenhouse conditions. Under field conditions, F2:3 progeny disease scores were skewed towards resistance, suggesting quantitative inheritance of resistance. In maize resistance to Turcicum leaf blight is both qualitative and quantitative. This study shows that resistance in sorghum to Turcicum leaf blight is quantitative suggesting that quantitative resistance in both maize and sorghum, close relatives, predates speciation. Breeding for such complex traits is often compounded by genotype by environment interactions and as such, marker assisted selection could hasten the process. Further characterisation of resistance loci and mapping of quantitative trait loci will support effective more resistance breeding. Key Words: Exserohilum turcicum, Sorghum bicolor, transgressive segregation, Uganda RÉSUMÉ Le sorgho (Sorghum bicolor L. Moench) a une importance socio-économique mondiale et est un modèle d’espèce botanique pour beaucoup d’herbes tropicales avec des génômes complexes. La culture est fréquemment dévastée par la maladie du flétrissement foliaire causée par Exserohilum turcicum avec de pertes considérables de rendements en grains et fourrage. Le développement des variétés résistantes à l’Exserohilum turcicum est le moyen le plus économique pour la gestion de la maladie. Par ailleurs, l’acquisition de la résitance au E. turticum dans le sorgho est mal comprise. Des recherches étaient conduites en Ouganda pour étudier le mode d’acquisition de la résistance au E.turticum dans le sorgho en serre et au champ. Des familles ségrégantes dérivant du croisement MUC007/009 (une accession locale résistante) et Epuripuri (une variété élite susceptible) étaient utilisées avec ces deux parents dans l’étude. Quatre témoins étaient aussi considérés lors des évaluations des familles, à savoir GAO6/106 (modérément résistant), Lulud (susceptible), MUC007/010 (résistant) et GAO6/18 (modérément susceptible). En serre, la sévérité de la maladie sur les plants F2 a montré une tendance asymétrique dénotant une consistante résistance. Au champ, la sévérité de la maladie était moins marquée sur le parent résistant que sur le parent susceptible. Par ailleurs, il n’y avait aucune différence entre les deux parents en conditions de serre. En champ, les scores des maladies sur F2:3 illustraient sur courbe une tendance à la résistance, suggérant une acquisition quantitative de la résistance. Dans la résistance du maïs au Turcicum, le flétrissement foliare est qualitatif aussi bien que quantitatif. Cette étude montre que la résistance au flétrissement foliare par Turcicum dans le sorgho est quantitave suggérant qu’une résistance quantitative dans le sorgho et le maïs prédate la spéciation. L’amélioration pour de tels traits complexes est souvent liée aux interactions génotype x environnement et ainsi, la sélection par marqueur assisté pourrait accélérer le processus. Plus de caractérisation et de cartographie des traits loci pourront permettre une amélioration effective de la résistance. Mots Clés: Exsherohilum turcicum, Sorghum bicolor, ségrégation transgressive, Ouganda INTRODUCTION Sorghum (Sorghum bicolor L. Moench) is a tropical C4 monocot plant and a subject of plant genomics research (Paterson, 2008). It has a relatively small genome of about 750 million base pairs (Arumuganathan and Earle, 1991). Also, it has a small amount of repetitive DNA and has co-linearity with other cereal genomes (Kong et al., 2000). Turcicum leaf blight caused Exserohilum turcicum (Pass) K.J. Leonard and E.G. Suggs (Teliomorph: Setosphaeria turcica [Luttrell] Leonard and Suggs) is an old disease of sorghum and maize (Agrios 1997; Ramathani et al., 2011). Under many tropical conditions and especially on susceptible genotypes, considerable grain and fodder yield losses of up to 50% can occur (Mittal and Boora, 2005; Ogliaril et al., 2007). Recent studies in Uganda show that cross infection of sorghum and maize is possible, with race 1, 2, 3 and 0 identified from sorghum isolated E. turcicum (Ramathani et al., 2011). Given that both maize and sorghum have high synteny with shared ancestry (Swigonová et al., 2004), it is possible that resistance to E. turcicum in both crops may predate speciation and be similar crops. The occurrence of cross infection of maize and sorghum supports this hypothesis. In maize, several dominant or partially dominant qualitative genes have been described that confer race-specific resistance, including Ht1 (Hooker, 1963), Ht 2 (Hooker, 1977), Ht 3 (Hooker, 1981), Htn 1 (also known as HtN, Gevers, 1975) and Ht P (Ogliari et al., 2005; Carson, 1995) A quantitative resistance system has also been reported (Hooker 1981: Adipala et al., 1993). Developing varieties with resistance against E. turcicum is the most cost-effective way to manage the disease. However there is paucity of information needed to support resistance breeding. The objective of this work was to determine the mode of inheritance of resistance to TLB on sorghum using disease response of segregating lines and using generation mean analysis. MATERIALS AND METHODS Description of research area. This study was carried out in Uganda at Makerere University Agricultural Research Institute Kabanyolo (MUARIK). The Institute is at an elevation of 1200 m above sea level (O°28’N and 32°37’E) (Tenywa et al., 2001). The populations used in the study were developed at the National Semi Arid Agricultural Research Institute (NaSAARI), Soroti and at MUARIK. Genetic materials and population characterisation. A total of 304 F2 segregating plants, and 278 F2:3 and 246 F2:4 segregating families were derived from a cross of MUC007/ 009 (resistant to leaf blight (Exserohilum turcicum)) and Epuripuri (elite but susceptible sorghum variety). Evaluations included the two parents and four checks GAO6/106 (moderately resistant), Lulud (susceptible), MUC007/010 (resistant) and GAO6/18 (moderately susceptible). No selection was made for resistance to TLB or for any agronomic traits during the development of the three populations. It should be noted that the number of plants in F2:4 populations reduced 19% because of germination failure and seedling death of some F2 and F2:3 individuals. Layout of the experiments. The three populations with their parents and the checks were evaluated for reaction to TLB at 51 days after planting (DAP). Greenhouse evaluation of the F2 in April to June of 2009 and field evaluation of the F2:3 population in October to December of 2009 were not replicated, while an alpha lattice design (5 blocks of 11 plots/block) of two replications was used in the field for the F2:4 Inheritance of resistance to Turcicum leaf blight in sorghum population in April to June of 2010. The F2:3 and F2:4 families were obtained from 3-5 representative plants from the previous generation, with equal portions of seed bulked for evaluation. Inoculum preparation and inoculation techniques. Infected sorghum leaves were collected from the field. Lesions were cut from these leaves and placed on moist paper towels in petri dishes for 48 hours to allow sporulation (Carson, 1995). Single spores were picked from the lesions and placed on potato dextrose agar (PDA) plates and incubated at room temperature in a dark place. Individual spores of E. turcicum were subsequently sub-cultured to fresh PDA plates used to inoculate autoclaved sorghum kernels, and allowed to colonize the sorghum kernels for about 14 days before use as source of innoculum (Carson, 1995). The colonised sorghum kernels were air-dried prior to field inoculation. Inoculation was done at the five leaf stage (Stage 2, Vanderlip, 1993) by placing 20 to 30 colonized sorghum kernels into the leaf whorls. Inoculation was done in the evening when dew and ambient temperature were optimal for successful infection (Carson, 1995). Disease assessment and evaluation and data collection and analysis. Disease severity was assessed using a scale of 0 to 75 where 0= no lesions identifiable on any of the leaves and 75 = 45 - 75% of leaf surface diseased (Adipala et al., 1993). Assessment commenced at stage 4 (the growing point differentiation) (Vanderlip, 1993). Weekly assessments of disease severity were used to compute area under disease progress curves (AUPDC) as described by Campbell and Madden (1990) and Adipala et al. (1993). The AUDPC values were standardized relative AUDPC by dividing the AUDPC, values total duration of the of epidemics (Madden et al., 2008). All data were subjected to analysis using GenStat Discovery Edition 12 (LAWES Agric. Trust, 2010). RESULTS Disease reactions to Turcicum leaf blight in F2 , F2:3 and F2:4 populations. The AUDPC in the greenhouse were skewed towards resistance, with a mean AUPDC of 11.01 (Table 1) (Fig.1). Out of 304 F2 progeny, 194 had severity scores of less than 12% percent leaf area affected. Under greenhouse conditions the resistant parent had relatively high severity score compared to another resistant check MUC007/ 010 (AUDPC 5.8) and the moderately resistant genotype GAO6/106 (AUDPC 23.65 (Table 1). Lulud (susceptible check) had the highest AUDPC of 36.7 and the moderately susceptible GAO6/18 had exceptionally high AUDPC of 98.7 compared to the susceptible parent Epuripuri (AUDPC 29.4) (Table 1). The unpredictable responses of the known genotypes prevented any clear interpretation of the response of the segregating progeny. In the field environment, F2:3 disease severity scores were skewed towards resistance indicating the possibility of partial dominance conditioning resistance in sorghum to TLB (Fig. 2). The F2:3 mean AUDPC was 12.1, almost identical to the mid-parent value AUDPC was 13.9. The moderate resistant check variety GAO6/106 had AUDPC ratings of 3.4 similar to that of the resistant parent MUC007/009 4.0 and to the resistant check MUC007/010 (4.5) (Table 1). The susceptible check GAO6/18 had surprisingly low disease severity (7.1), than MUC007/009 (5.9) (Table 1). Under the field environment the mean AUDPC values of F2:4 segregating families was 6.6, equal to the mid-parent value (Table 2). The resistant parent (MUC007/009) severity rating was 4.6 while the susceptible parent (Epuripuri) rating was 8.6. AUPDC and disease severity ratings of F2:4 segregating families were significantly different between families (Table 2). Due to loss of about 19% of F2:4 families, (susceptible and resistant families), well-defined interpretation of the segregating patterns of these families was difficult (Data not shown). The moderately resistant genotypes GAO6/106 and GA06/18 had similar severity ratings of 5.5% and 5.2%, respectively, which were just slightly higher than the resistant parent MUC007/009 (4.61) (Table 2). DISCUSSION Disease severity of F2 plants in the greenhouse and F2:3 families in the field were skewed towards resistance, suggesting quantitative inheritance with mainly additive effects but with partial dominance toward resistance. The resistant parent had much lower disease severity than the susceptible parent under field conditions. In maize resistance to TLB is conditioned by quantitative and qualitative mechanisms (Hooker, 1963; Gevers, 1975; Hooker, 1977; Hooker, 1981; Ogliari et al., 2005). In the E. turicum –maize pathosystem, quantitative resistance is characterized low lesion number, small lesion area with typical necrotic lesion types, as well as reduced severity and AUDPC values; whilst qualitative resistance is characterised by small lesions surrounded by chlorotic halo also referred to as Ht (Helminthosporium turcicum)-lesions type. This Ht lesions were first observed on lady finger popcorn in the 1940s and subsequently on other maize genotypes (Hooker, 1963, 1981; Gevers, 1975; Adipala et al., 1993). In this study, the resistant plant had reduced AUPDC values and fewer lesions which were generally small in size (data not shown), typical of quantitative resistance to TLB. Indeed, segregating patterns of were indicative of quantitative inheritance with possibility of partial dominance towards resistance. Quantitative resistance in general, supports mild levels of epidemics, invariably allowing the existence of a variable population of pathogens (McDonald and Linde, 2002). In the Uganda, and many other tropical countries were E. Turicum has several alternative hosts which support pathosystem robustness for both resistance (on the part of host) and virulence (on the part of the pathogen) (Chiang et al., 1989), there is need for mechanisms such as quantitative resistance which do not create extreme selection pressure and speed up patho- evolution (Dangle and Jones, 2001; McDonald and Linde, 2002). Moreover, the occurrence of quantitative resistance as the basic mechanism to protect the plant is a logical consequence given that sorghum was domesticated in Africa. Whereas the Ht based resistance is well characterised in maize, and shows dominance (Hooker, 1963; Gevers, 1975; Hooker, 1977; Hooker, 1981; Ogliari et al., 2005) in this study we did not find evidence of this type of dominant gene action. Breeding for quantitative resistance is albeit challenging often due to compounding effects of genotype by environment interactions that can be compounded by a variable pathogen population. In this study this was the case, the greenhouse had higher severity than the field. TLB disease epidemics are favoured by high rainfall and relative humidity and moderate temperatures (Hennessy et al., 1990; Carson 2005), so higher disease severities in the greenhouse than in both field evaluations were not surprising, since E. turcicum is a necrotroph that requires high humidity and warm temperature for infection (Adipala et al., 1993); and such conditions are common under greenhouse conditions. Taken together, the results of this study suggest that resistance in sorghum to E. turcicum is quantitative, with some contribution of additive, dominance and epistatic effects. Results also highlight that environment can have major effects on the disease response of specific sorghum genotypes to Turcicum leaf blight. ACKNOWLEDGMENT We would like to extend our sincere gratitude and appreciation to the Regional Universities Forum for Capacity Building in Agriculture (RUFORUM), the Programme on Strengthening Capacity for Agricultural Research and Development in Africa (SCARDA) for support while undertaking this research, the manuscript was written with the support of Professor P. Gibson. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12022f1.jpg] [cs12022t1.jpg] [cs12022t2.jpg] [cs12022f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}