 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
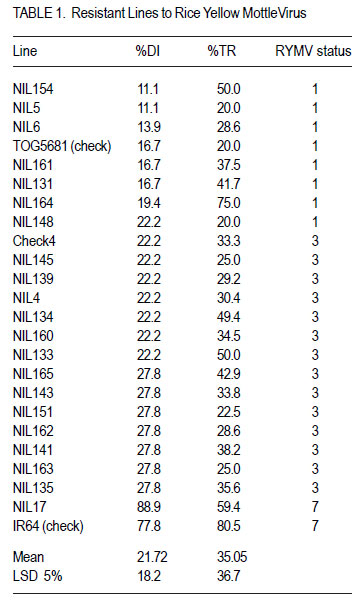
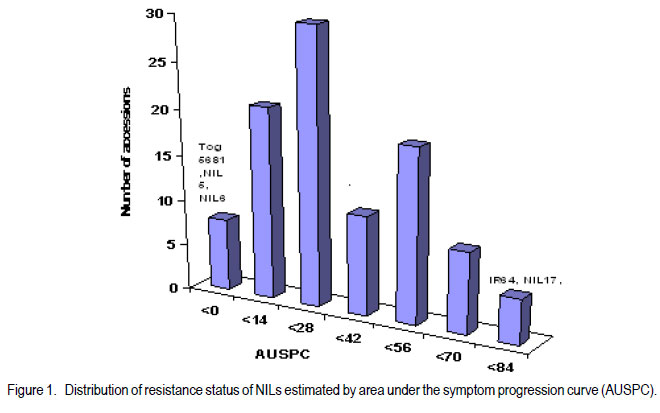
African Crop Science Journal, Vol. 20, Supp. 1, 2012, pp. 163-168 Identification of near-isogenic lines resistance to rice yellow mottle virus A. Jaw, M.N. Ndjionndjop1, R. Akromah and Y. Séré1 Kwame Nkrumah University of Science and Technology, Kumasi, Ghana Code Number: cs12023 ABSTRACT Rice yellow mottle virus (RYMV) is a serious disease in rice production in the lowland and irrigated ecologies. A study was conducted to examine the resistance of rice ( Oryza sp.) lines RYMV. One hundred near-isogenic lines (NILs) from BC2F7 population derived from cross combination: Gigante (Oryza indica cv.) x IR64, Gigante x FK28 and Gigante x IR47 were evaluated. Twenty NILs were identified to be resistant to RYMV BF27 isolate from phenotypic screening. Enzyme-Linked-immunosorbent assay (ELISA) test revealed 36 lines to be resistant to RYMV with low virus content. Foreground selection using the gene marker, revealed 22 lines showed introgression of rymv1-2 allele. On the average, 71% of the markers used in the evaluation showed polymorphism. The highest proportion of recurrent parental contribution was obtained from the cross Gigante x IR64; and the lowest from Gigante x IR47. Individuals from Gigante x FK28 had the highest percentage of the donor parent (70%) and Gigante x IR64 had the lowest value but showed the highest genomic proportion of the recurrent parent (57%) was showed by Gigante x IR64. Key Words: ELISA, polymorphism, Oryza spp. RÉSUMÉ Le virus de la panachure jaune est une maladie grave pour la production du riz ( Oryza sp.), spécialement le riz des bas-fonds et des écologies irriguées. Une étude était menée pour examiner la résistance des lignées de riz au virus de la panachure jaune (RYMV). Une centaine de lignées quasi-isogéniques (LQI) provenant d’une population de rétrocroisement BC2F7 issue de la combinaison des croisements: Gigante (Oryza indica cv.) x IR64, Gigante x FK28 et Gigante x IR47 étaient évaluées. Vingt LQI étaient jugées résistantes à l’isolat RYMVBF2F7 sur base de la sélection phénotypique. Le test ELISA montrait 36 lignées résistantes au virus de la panachure jaune avec un faible contenu en virus. La sélection de premier plan basée sur le marqueur génique montrait que 22 lignées manifestaient l’introgression de l’allèle rymv1-2. En moyenne, 71% des marqueurs utilisés manifestaient un polymorphisme. La plus grande proportion de la contribution de la part du parent récurrent était obtenue du croisement Gigante x IR47. L’individu provenant du croisement Gigante x FK28 avait le plus grand pourcentage du parent donneur (70%) et le croisement Gigante x IR64 avait la plus faible valeur, mais montrait que la plus grande proportion génomique du parent récurrent (57%) était manifestée par Gigante x IR64. Mots Clés: ELISA, polymorphisme, Oryza spp. INTRODUCTION Rice ( Oryza sp.) is one of the main staple food crops in many parts of Africa and it accounts for 20 -50% of total caloric consumption in the bulk of the world populations (Nutsugha et al., 2004). Despite the increased area under rice cultivation, yields remain far below the level of production, and is largely in the range of 1 to 3 t ha-1 (Traoré et al., 2006). Optimum rice production in Africa cannot be realised mainly due to prevalence of diseases. Rice yellow mottle virus (RYMV) is among the most devastating and yield reducing diseases. It was first discovered in Kenya in the 1960s (Bakker, 1974), and has remained one of the most problematic diseases in Africa. Rice yellow mottle virus is believed to be transmitted by beetles (Bakker, 1974), but also by animals like cattle, donkeys and rats; and through wind (Sarra and Peters, 2003; Sarra et al., 2004). Rice cultivars, especially those of the Oryza sativa indica are susceptible to RYMV; and they suffer yield losses of up to 100% (Abo et al., 1998). However, Oryza sativa, Gigante and some traditional rice cultivars (Oryza glaberrima) Tog 5681; Tog 5672; Tog 5675 and Bekarosaka are highly resistant to RYMV (Ndjiondjop et al., 1999). High resistance behaves as a monogenic recessive genetic trait and is associated with lack of symptom development and restriction of virus movement (Ndjiondjop et al., 2001). Fortunately, with genetic map of molecular markers, the high level of natural resistance has facilitated the introgression of the resistance gene into popular RYMV susceptible elite varieties using markerassisted selection . During the past years, several near-isogenic lines with rymv1-2 resistant allele were developed by the Biotechnology Unit at Africa Rice Center, and have been evaluated for resistance to rymv in the Republics of Mali and Guinea (Dr. Ndjiondjop, Africa Rice Center, Benin, Pers. com.). One hundred lines were selected based on level of resistance. The objective of this research was to validate and confirm the resistance of nearisogenic lines obtained from the introgression of rymv1-2 gene from Gigante into locally-adapted but susceptible varieties. MATERIALS AND METHODS A total of 100 near-isogenic lines (BC2F7), 7 parental lines and 3 checks were evaluated in this study. Seeds were sown in 1-litre pots at a rate of six seeds per pot and then thinned to three seedlings. Virus source and inoculum preparation. The most aggressive isolate B27 originated from Northern part of Benin (Dr. Séré, Africa Rice Center, Benin, Pers. com.). In order to get sufficient material, the isolate was multiplied on the standard susceptible variety IR64, and was recovered by mechanical inoculation of the susceptible cultivar by rubbing sap extract on the upper and lower leaf surface of 3 weeks-old seedlings. This was done with the aid of carborundum. Leaves that exhibited symptoms of the disease at 21 days after inoculation were harvested to prepare sap inoculum source for evaluation. Infected leaf samples were ground in distilled water (1g of infected leaf to 10ml of distilled water) using a sterile blender. Carborundum was added to the extracts which were subsequently rubbed to whole leaves of 21 days old seedling, from the base to the top with the fingers dipped in the viral sap. Visual scoring. Phenotypic screening was carried out in a screenhouse at the Africa Rice Center in Cotonou, Benin. A hundred nearisogenic lines, 7 parental lines and 3 controls were evaluated with the most aggressive local isolate BF27 (S1 strain) to prevent introduction of another isolate. An augmented randomised experimental design was used. Observations of the symptoms were recorded weekly, up to 42 days after inoculation. A 1 to 9 symptom scoring scale using IIRI standard evaluation system was used: where 1 scored for no symptom observed, 3 for leaves green with sparse dots streak, 5 scored for leaves pale green, 7 scored for pale yellow leaves or yellow, mottling with reduction in height and 9 scored for leaves yellow or orange with necrosis and sometimes plain death. Chlorophyll content and agronomic traits (height and tiller number) were recorded at 28 and 42 days after inoculation. Distribution of the area under the symptom progressive curve (AUSPC) was calculated using the formula proposed by Thiémélé et al. (2010); AUSPC = [Si +S1+i) -2) (ti+1)-t1)]/2; where Si corresponds to the symptom at date ti in days) using the two dates (28 and 42 DAI). Inoculated and infected systemic leaf samples at 42 days after inoculation were collected for enzyme-linked immunosorbent assay (ELISA) test to check for the virus content using Antigen Coated-plate ELISA (ACP-ELISA). Inoculated and systematic infected leaf samples were ground in coating buffer (Na2 CO3, NaHCO3, pH 9.6) at 1:10 (0.1 g to 1000 µl) dilution. A sample of 200 µl virus sap extract was added in duplicated wells of microtitre plate and was incubated overnight. After overnight incubated, plates were washed with PBS-T buffer 4 times. Two hundred microlitres (200 µl) of 1% bovine serum albumin (BSA) was used to block each well and incubated at 37 °C for 1 hour. After incubation, the solution (BSA) was poured off without washing. Polyclonal antibody 1:1000 in phosphate buffer saline, was prepared and wells were blocked with 200 µl and incubated at 37 °C for 2 hours. After incubation for 2 hours at 37 °C, wells were blocked with 200 µl of goat anti-rabbit alkaline phosphate (Sigma) conjugate to 1:1000 in conjugate buffer (PBS + 2% PVP-40 polyvinyl pyrrolidine + 0.2% egg albumin (Sigma A-5253) was added to each well and incubated at 4 °C over night. After overnight incubation at 4 °C 200 µl of freshly prepared substrate (1mg ml-1 of p-nitrophenyl phosphate [Sigma] in substrate buffer (diethanolamine, sodium azide (NaN3, H2O) pH 9.8) was added to each well and incubated at room temperature for 1 hour. Results were assessed by visual observation and reading of samples with spectrophotometer at 405 nm. Disease progression was also calculated. Data collected were analysed using IRRISTAT version 5.0 software. Molecular genotyping. Genomic DNA was extracted from 3-weeks old seedlings using Mixed Alkyltrimethyl-ammonium Bromide (MATAB) adapted from CYMMYT protocol. About 36 nearisogenic lines identified from the phenotypic screening were used for the molecular characterisation. Ten seeds were planted per line and bulk-harvested for genomic DNA extraction. The isolated genomic DNA was dissolved in sterile distilled water. To estimate the DNA concentration, a spectrophotometer was used and the quality was checked by electrophoresis in 1% agarose gel, and detected by staining with ethidium bromide. Polymerase chain reactions were carried out in a final volume of 25 µl which consist of 30 ng DNA, 10X PCR buffer (100 mM Tris-HCl, MgCl2 15 mM, 500 mM KCl, 1% triton X 100) 0.1 mM of each dNTPs and 10 µM of each of the forward and reverse primers. The amplification was carried out in a thermocycler consisting of an initial denaturation at 94 °C for 4 minutes, followed by 30 cycles of denaturation at 94 °C for 30 seconds, annealing at 55 °C for 30 seconds, extension at 72 °C for 45 seconds and final extension at 72 °C for 5 minutes. The amplification was separated on a 3% agarose gel and stained with ethidium bromide. One hundred and twenty (120) microsatellites or simple sequence repeat primers were used to screen for polymorphism. Data scoring and analysis. The marker RM252 which is linked to the resistant gene of rymv1-2 was used to check for the presence of the resistance gene. The introgression of the resistance gene was assessed by the banding patterns obtained from the amplification. The lines that carried the introgressed resistant gene of Gigante were further genotyped to check their genomic composition of the recurrent parents. Polymorphic markers were used to check the background of the lines resistant to RYMV. Bands were scored as A for donor parent allele, B for the recurrent parent allele, H for the heterozygous allele and U for the non-parental allele. Data were analysed using graphical genotype (GGT 2.0) software package (Van Berloo, 1999). RESULTS Phenotypic screening. The screening work investigated the resistance of near-isogenic lines to RYMV by visual screening and was confirmed by ELISA test. About 91 of the lines germinated out of 110 and analysis was based on these numbers of lines. On an average, the effect of RYMV appeared at 28 and 42 days after inoculation. Therefore, analysis was done on these two dates. Based on disease incidence, there was a significant difference between lines (P < 0.01) as well as days after inoculation (P < 0.01). Analysis of variance showed significant differences in height reduction between lines (P < 0.01). However, no significant difference was observed between the days after inoculation (P > 0.223). There was a significant reduction in tiller number reduction among the lines (P < 0.01). The interaction between varieties and days after inoculation also showed significant differences in tiller number reduction (P < 0.01). Results from the visual expression can be grouped into 3 categories. The first group showed lines resistant to BF27 with disease incidence ranging from 11.1 to 27.8 % (Table 1). The second group showed moderate resistance with disease incidence ranging from 33.3 to 50%; and the third group showed susceptibility with disease incidence ranging from 55.6 to 88.9%; with clearly visible symptoms, yellowing and mottling. The resistant check (Tog 5681) was classified in the resistant group; whereas Gigante was in the moderately resistant group. Among the nearisogenic lines screened, 24% showed resistance, 46% moderately resistance and 30% susceptible. The distribution of the area under the symptom progressive curve (AUSPC) ranged from 0 to 84, with different frequencies (Fig. 1). The most susceptible line started to develop symptoms before 7 days after inoculation, like the susceptible check (IR64). This was accompanied by reduction in growth. About 8 of the lines had no symptom progress within the two dates with AUSPC value of zero. Twentyone of the lines presented mild symptoms with disease progress scored from 1 to 3, and with AUSPC value of 14 within the two dates. Some of the NILs and IR64 which were susceptible, had high values ranging from 70 to 84 (Fig. 1). Results from the ELISA test revealed 36 lines to be resistant and 22 to be moderately resistant. The level of virus content ranged from 0.6 to 4.9%. Genotyping. The lines which were identified from the ELISA test were used to check the introgressed gene resistance to RYMV. The foreground of these lines was identified using the marker linked to the gene rymv1-2. About 22 of the NILs showed the resistance gene rymv1-2 of Gigante. Polymorphism evaluation revealed 40 to 73% of the markers to be polymorphic between Gigante and the other parental lines. Polymorphic markers were used to determine the genomic proportion of resistant NILs. Results indicated that the markers used between Gigante and IR64 showed 57% of the IR64 allele, 34% of the Gigante allele and 9% showed non-parental allele (U). For Gigante and FK28, 19% of the FK28 allele, 70% of Gigante allele, 9% non-parental lines and 2% heterozygote. DISCUSSION There was a response difference of lines resistant to RYMV, (Table 1) some of the lines were not resistant to RYMV. These results are supported by Kouassi et al. (2005) who reported that the responses to RYMV depends on the genotype, screening condition, severity of the inoculation and the evaluation method. Early symptoms were noted at 7 DAI in IR64 (susceptible check) and some of the NILs and this is consistent with early symptom development on susceptible cultivars such as IR64, Tog5673 (Ndjiondjop et al., 1999). Although field test of these near-isogenic lines reported the resistance of these lines, this research results showed that natural field screening is not the only method that can discriminate or confirm the resistance to RYMV. Mechanical inoculation under a controlled environment could be a better discrimination procedure. In addition, this research is consistent with early reports which stated that durability of resistance to RYMV should not be assessed by natural existing isolates, but by mechanical inoculation of commonly existing isolates (Fargette et al., 2002). Traoré et al. (2006) reported that 16.4% of some isolates can overcome the resistance of Gigante, recent reports pointed out that only small proportion of isolates have the ability (Poulicard et al., 2009). At 42 DAI, IR 64 and NIL17 followed the same trend: plants become stunted, yellowing and necrosis was noted. This reveals that some NILs have the same degree of susceptibility like IR64. ELISA test confirmed 36 lines to be resistance to RYMV. This indicated that symptom assessment in screening test is not enough to discriminate lines because symptoms can be masked in some cultivars and with some isolate (N’Guessen et al., 2001). The introgressed resistance gene of rymv 1-2 was identified in 22 NILs (Table 1). These results highlighed the importance of marker-assisted selection in identifying a targeted gene in which direct selection is difficult or impossible in breeding programmes. In this study, a total of 120 SSR markers were used for the polymorphism test. On an average, 71% of the SSR markers showed polymorphism among parental lines. Based on the data obtained from the polymorphic markers, the proportion of each parental line varied among the individuals. Babu et al. (2004) reported that recurrent backcrossing programmes are planned on the assumption that the proportion of recurrent parent genome is recovered at a rate of 1- (1/2) t+1 for each of t generation of backcrossing. In this study the average proportion of recurrent parent DNA in the lines was less than what is expected for a random set of BC2 lines (87.5%) and the donor parent (12.5%) (Semagn et al., 2007). Based on the results obtained, this study suggests that additional backcrossing is needed in order to improve the genomic proportion of the recurrent parents in these nearisogenic lines. CONCLUSION Out of the 100 near-isogenic lines, phenotypic screening revealed 20 of the lines expressed resistance and 22 of the lines manifested the resistant gene of Gigante. Although the management of rice yellow mottle virus using cultural practices help to reduce the impact , the use of varietal resistance is more durable. In addition, the integration of screenhouse experiment together with marker-assisted selection will enhance the ability of the breeders to determine the level of inheritance and know the introgressed gene from parents to offspring. ACKNOWLEDGMENT The study was financed by the Strengthening Capacity for Agricultural Research and Development in Africa (SCARDA), networking project and the Biotechnology Unit of Africa Rice Center. REFERENCES
Copyright © 2012 - African Crop Science Society The following images related to this document are available:Photo images[cs12023t1.jpg] [cs12023f1.jpg] |
| |||||||||

{kind=link}
{kind=link}