 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
CROP TO WEED GENE FLOW IN SORGHUM: IMPLICATIONS FOR TRANSGENIC RELEASE IN AFRICA P.E. ARRIOLA Department of Botany & Plant Sciences, University of California, Riverside, CA 92521, USA
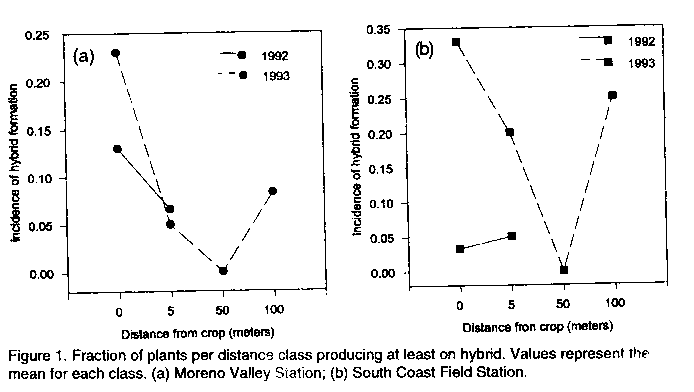
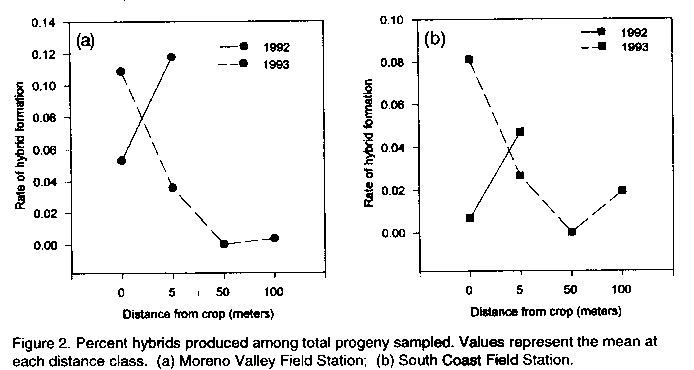
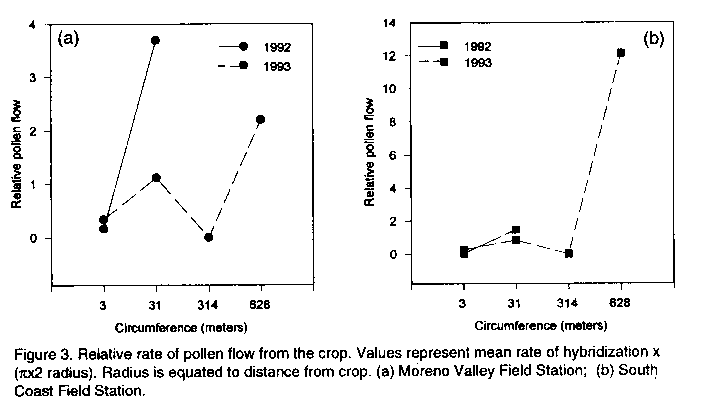
Code Number: CS95021 Size of Files: Text: 27K Graphics: Line Drawings (gif) - 22K ABSTRACT Potential problems with the release of transgenic crops are varied. Among these problems is the threat of gene flow via pollen transfer from the genetically altered crop to wild or weedy plant relatives growing nearby. Because some engineered genes may confer an adaptive advantage, troublesome weeds may become more problematic if they bear such transgenes. Although the rate of crop to weed gene flow has been measured for a few systems, large gaps in our understanding of this phenomenon remain. The likelihood of crop to weed gene flow in a crop/weed system involving the crop, Sorghum bicolor, and a related weed, S. halepense, in North America is described. Measurements of the rate and incidence of hybridization were made using progeny analysis. Spontaneous hybridization under field conditions was detected at distances of 100 metres. Measurements of several fitness correlates were taken on hybrid weeds and compared to non-hybrid weeds in order to address the potential for persistence of transgenes in the environment. No significant differences were recorded between hybrid and non-hybrid weeds, suggesting that beneficial genes introduced into weed populations may persist. The implications for the release of transgenic sorghums in Africa are discussed. Key Words: Pollen transfer, Sorghum bicolor, S. halepense, transgenes, weeds RESUME Les problemes potenfiels lies a la diffusion des cultures transgeniques sont varies. Parmi ces problemes il ya a la menace de la dispersion de genes par le transfert de pollen de la culture genetiquement alteree aux parents sauvages ou herbaces qui poussent dans les environs. Comme certains genes peuvent conrefer une adaptation plus avantageuse, les mauvaises herbes qui les acquierent peuvent devenir plus problematiques. Bien que le taux de dispersion de genes de culture vers les mauvaises herbes ait ete mesure dans le cas de quelques systemes, la comprehension de ce phenomene n'est pas encore totale. Le papier dectit le passage de gene d'une culture telle que le sorgho vers une mauvaise herbe, S. halepense, au nord de l'Amerique. En examinant les progenitures, il etait possible de mesurer le taux et l'incidence de l'hybridation. L'hybridation spontanee en conditions de champ etait detectee a 100 metres de distance. Les mesures de plusieurs correlations etaient faites sur les mauvaises herbes hybrides et comparees aux non-hybrides en vue de determiner la persistance de genes transferes dans cet environnement. Il n'y avait pas de differences significatives entre hybrides et non-hybrides de mauvaises herbes; ceci indique que les genes benefiques introduits dans les populations de mauvaises herbes pourraient persister. Le papier discute aussi les implications dues a la dissemination des sorgho transgeniques en Afrique. Mots Cles: Transfer de pollen, Sorghum bicolor, S. halepense, transgenes, mauvaises herbes INTRODUCTION Gene flow is the movement and incorporation of genes from the gene pool of one population to the gene pool of another population (Slatkin, 1987). Substantial gene flow has profound effects on the genetic structure of populations (Ellstrand, 1992), leading to changes in their composition, to adaptive shifts, or even to the extinction of some populations (Small, 1984; Futuyma, 1986; Slatkin, 1987). Crop plants which are sown near populations of wild or weedy relatives are often capable of gene exchange with those wild or weedy plants (Baker, 1972, 1974; de Wet and Harlan, 1975). A small but growing set of experimental data indicates that gene flow events between crops and weeds may be more common than previously believed (Klinger et al., 1991, 1992; Till-Bottraud et al., 1992; Arias and Rieseberg, 1994; Wilson and Manhart, 1994). The consequences of crop/weed mating events largely depend on the direction of pollen transfer. Effects can be varied, ranging from breeding progress retardation (Smith and Adams, 1983), to apparent fitness boosts in the crop/weed hybrids (Klinger and Elistrand, 1994), to crop improvement (Prescott-Allen, 1983; Ladizinsky, 1992). However, the likelihood and consequences of crop/weed matings on a large scale remains generally unknown for most crops. Most research has been focused on gene flow in the weed to crop direction. Hybridization in the crop to weed direction has received limited attention and its significance remains controversial. The National Research Council (1989) states that crop to weed hybridization should be of little concern because hybrid weeds which carry traits deemed desirable in crops (i.e. dwarfing or non-shattering seeds) would be at a competitive disadvantage with the weeds which do not carry such traits, therefore crop genes should not persist in weed populations. However, it has also been argued that crop to weed hybridization can and does lead to the enhancement and continued evolution of weedy plant pests (Baker, 1974; deWet and Harlan, 1975; Barrett, 1983). This controversy has particular contemporary significance. The advent of new biotechnologies have allowed the genetic transformation of crop plants for specific traits, many of which may confer an adaptive or competitive advantage to the weedy plants which acquire them (Ellstrand and Hoffman, 1990; Raybould and Gray, 1994). Scientists must now address the environmental consequences of the escape of transgenes into wild or weedy plant populations. The focus of the research below is to shed light on the occurrence of crop to weed mating in the genus Sorghum, the consequences of such matings, and the implications of crop to weed gene flow for the planned introductions of genetically modified crops into Africa. Experimental systems addressing crop to weed hybridization include: Brassica (Eber et al. 1994; Jorgensen and Andersen, 1994), Otyza (Langevin et al., 1990), Beta (Santoni and Berville, 1992; Boudry et al., 1993; ), Cucurbita (Kirkpatrick and Wilson, 1988), Gossypium (Umbeck et al., 1991 ), Raphanus (Klinger et al., 1991,1992), Setaria (Till-Bottraud et al., 1992), Chenopodium (Wilson and Manhart, 1994) and Helianthus (Arias and Rieseberg, 1994). Such studies have shown that crop to weed gene flow varies with pollen vector, weed distance from the crop, weed relatedness to the crop, and the level of precaution taken to minimize pollen flow from the crop: .Summarized below are the results of a study (Arriqla, 1995) that examines the rate and incidence of crop to weed hybridization in a system involving crop sorghum, Sorghum bicolor (L.) Moench, and a related weedy plant, S. halepense (L.) Pers., and the fitness of their hybrids. CROP TO WEED HYBRIDIZATION IN SORGHUM Sorghum bicolor (L.) Moench is one of the most economically important crops in the world with over 40 million hectares dedicated to its cultivation (FAO, 1993). Its many uses have made it a staple of world-wide agriculture for years. Its economic importance world-wide has also made it a target for improvement via genetic transformation. For example, S. bicolor has recently been transformed with a herbicide resistance marker (Casas et al., 1993). A wind pollinated annual, S. bicoloris sympatric and cross compatible with several wild congeners (Doggert, 1988). Circumstantial evidence suggests that the crop spontaneously hybridizes with Sorghum halepense (L.) Pers,johnsongrass, under field conditions in the United States (Baker, 1972). Johnsongrass is a wind pollinated, tetraploid (2n=40) perennial which reproduces vegetatively as well as through sexual means. It is among the world's ten worst weed pests (Holm et al., 1977). Johnsongrass is primarily a self-pollinated plant but will outcross ( Warwick and Black, 1983; Kigel and Rubin, 1985). Crop sorghum and johnsongrass occur sympatrically virtually everywhere the crop is cultivated, and the flowering times of the two plants overlap (Holm et al., 1977). Forced-hand crosses of S. bicolor x S. halepense have been successful under greenhouse conditions (Sangduen and Hanna, 1984). In order to estimate the likelihood of gene transfer from crop sorghum to its related weed, johnsongrass, an experimental crop weed system was set up. Dwarf milo sorghum was sown in a 0.2 ha stand simulating typical commercial cultivation at two locations in California over two consecutive years. Johnsongrass was planted at varying distances (0.5, 5, 50, and 100 m) around the crop to simulate a weedy infestation of the area surrounding the field. The crop and weed were of known genotypes, fixed homozygotes for alternate alleles at a single isozyme locus. Rate and incidence of hybrid formation were measured using progeny analysis. Also, forced-hand crosses in the crop to weed direction were made in order to generate F, hybrids for a comparison of several fitness correlates between hybrid and non-hybrid weeds. Percentages of hybrid progeny among seedlings germinated in the first year after exposure to the crop were extremely variable. Hybrid offspring accounted for 0-100% of the germinated seedlings on a per plant basis. Incidence of hybridization, (proportion of targets producing at least one hybrid seed), tended to decline with increasing distance from the crop (Figures 1a, b). Hybrids were detected at distances of up to 100 m from the crop. The 0.5 m distance values at the South Coast Field Station between 1992 and 1993 for incidence were the only ones found to differ significantly P < 0.05). Rate of hybridization, (proportion of the total progeny sampled which were hybrids), also tended to decline with increasing distance from the crop (Figures 2a, b). Differences in the total rate of hybrid formation between locations within years were significant (P< 0.05); however, no other significant comparisons were made. Estimates of relative pollen dispersal were made from the hybridization rates by multiplying them by (pi) d (diameter) (Figure 3a, b). Calculated values are estimates of relative pollen flow required for observed rates of hybridization. Relative pollen flow required at 0.5 m was lower than that at 100 m for hybrid formation.
Figure 3. Relative rate of pollen flow from the crop. Values represent mean rate of hybridization x (pi x 2 radius). Radius is equated to distance from crop. (a) Moreno Valley Field Station; (b) South Coast Field Station.
To determine if a crop can hybridize with a wild or weedy relative, however, is only the first part of meaningful risk assessment. One must determine the possible fate of the transgene in the environment. To address this, johnsongrass and sorghum were crossed in the greenhouse and F1 hybrids were produced. Several fitness correlates were measured in the hybrid and non-hybrid progeny in a common field plot. The characters measured included inflorescence production, days to first flower, tiller production, and pollen stainability. No significant differences in the measured values were found. However, hybrid weeds tended to have a higher pollen stainability than the non-hybrid weeds (P< 0.05). This observation suggests that any crop genes introduced into weed populations which are either neutral or beneficial are likely to persist. IMPLICATIONS FOR TRANSGENIC RELEASE There are many conditions which must be met if successful crop to weed hybridization is to occur. Four key conditions are (1) the crop and related wild or weedy plants must co-occur; (2) the crop and weed must be sexually compatible; (3) the crop and weed must have overlapping flowering times; and (4) there must be a pollen vector which can transfer the pollen from the crop to the weed. In the case of Sorghum, these conditions will almost certainly be met everywhere the crop is cultivated. Crop sorghum and its congeners are very often sympatric in their respective distributions. Johnsongrass affinity for Mediterranean climate and vigorous sexual and vegetative capacity for spread have enabled this plant to invade many areas of sorghum cultivation (Holm et al., 1977). In Africa, the centre of diversity for the genus Sorghum, there are a number of compatible members of the genus which are compatible with modern cultivated sorghum (de Wet and Harlan, 1971; Doggett, 1988). Introduction of modern cultivars into areas of plant diversity may pose a significant threat to wild landraces which occur there. Large stands of outcrossing modern cultivars could hybridize with the wild plants, driving them to extinction or homogenizing the gene pool so much that valuable genes could be lost (Ellstrand, 1992). The promiscuity of S. bicolor with the other members of the genus, such as S. propinquum, S. arundinaceum and S. sudanense. is well documented. Crop sorghum can and does readily hybridize with these sympatric relatives (Doggett, 1988). The flowering times of crop sorghum and its wild and weedy relatives overlap extensively. (Eastin and Lee. 1985; Doggert, 1988). Crop sorghum and its relatives are wind pollinated. The potential for spontaneous gene exchange in the field is real and is probably a common occurrence among the sorghums. Extensive and specific precautions, such as increased crop isolation distances. must be in place to minimize the risk of hybridization with transgenic plants. Opportunity for hybridization does not necessitate escape of transgenes. Several factors must be considered in the accurate evaluation of risk. For example. one must determine first whether a weed x crop hybrid will persist (Dale, 1994). The present results show that weed x crop hybrids of crop sorghum andjohnsongrass do not suffer any apparent fitness decreases when compared to non-hybrid weeds. This finding may be due to the fact that characters which have been selectively bred for in-crop sorghum may not contribute any detriment to the weed under cropping conditions. A field study under non- agricultural conditions where there may be more limiting factors might uncover measurable differences in hybrid fitness. However. in this study. a transgene would be expected to persist. A second issue which must be considered when estimating the possible persistence of an escaped transgene is whether there is a cost to the weed possessing the trait. Certainly ~he presence of a stress agent would favour retention and spread of the trait; for example, herbicide resistance would be expected to persist in weed populations if the pesticide were in use. However, the cost of such a gene might be too great in the absence of the selection pressure. In the latter environment, the hybrid weeds may not be as fit or as competitive as the non-hybrid weeds and thus the escaped gene might not persist. However, Gliddon (1994) points out that even if the hybrid weeds are not as fit as the non-hybrids, the transgene may persist and even increase in frequency if the magnitude of the gene flow from the crop is great enough to overcome the fitness decrease. The third case to be considered is that the transgene might provide no immediate benefit or cost and thus be considered neutral. Such a gene would be expected to persist in weed populations under continued gene flow until a future time when it might become a benefit or a liability. It is virtually impossible to determine the outcome in any specific case a priori. The only way we have to evaluate the true risk of transgenic release is through extensive field testing. The potential problem with escaped transgenes is directly associated with economic importance of the crops possessing them. The total hectarage dedicated to the cultivation of the top ten most economically important crop plants is immense. Thus, one can surmise, the greater the sown hectarage the more likely there will be a wild or weedy relative growing sympatrically with the crop. Many issues must be considered to suggest guidelines or recommendations for transgenic plant release. Hybridization and fitness experiments on a single group of plants are insufficient to make meaningful assessments of the true risk of transgenic release. Hybridization and fitness experiments need to be performed with weeds of many different genotypes from many different populations where the transgenic plant will be released. Performance of crop/weed hybrids which actually possess a transgene need to be compared to that of non-hybrid weeds under field conditions on a crop by crop basis. Certainly, in the case of Sorghum in Africa, the potential risk of gene escape warrants the strictest guidelines for transgenic release. ACKNOWLEDGMENTS I am grateful to N.C. Elistrand for his comments on the previous draft of this manuscript as well as for all of the help, support and guidance. I also thank J.M. Clegg, C.D. Arriola and D.R. Elam for the assistance on this project, and to the UC Riverside Graduate Affirmative Action Programme, Office of the Graduate Dean, and the Office of the President of the University of California for funding. The research was supported in part by a grant from USDA-CSRS #94-331200372 to NCE. REFERENCES Arias, D.M. and Rieseberg, L. 1994. Gene flow between cultivated and wild sunflowers. Theoretical Applied Genetics 89:655-660. Arriola, P.E. 1995. The Role of Crop to Weed Gene Flow in the Continued Evolution of a Noxious Weed, Sorghum halepense (L.) Pers. Ph.D. Dissertation, University of California. Riverside. CA, USA. Baker, H.G. 1972. Migrations of weeds. In: Taxonomy, Phytogeography, and Evolution. Valentine, D.H. (Ed.), pp. 327-347. Academic Press, London, U.K. Baker, H.G. 1974. The evolution of weeds. Annual Review of Ecological Systems 5:1-24. Barrett, S.C.H. 1983. Crop mimicry in weeds. Economic Botany 37:255-282. Boudry, P., Morchen, M., Saumitou-Laprade, P., Vernet, Ph. and Van Dijk, H. 1993. The origin and evolution of weed beets: consequences for the breeding and release of herbicide- resistant transgenic sugar beets. Theoretical Applied Genetics 87:471-478. Casas, A.M., Kononowicz, A.K., Zehr, U.B., Tomes, D.T., Axtell, J.D., Butler, L.G., Bressan, R.A. and Hasegawa, P.M. 1993. Transgenic sorghum plants via microprojectile bombardment. Proceedings of the National Academy of Sciences USA 90:11212-11216. Dale, P.J. 1994. The impact of hybrids between genetically modified crop plants and their related species: general considerations. Molecular Ecology 3:31-36. de Wet, J.M.J. and Harlan, J.R. 1971. The origin and domestication of Sorghum bicolor. Economic Botany 25: 128-135. de Wet, J.M.J. and Harlan, J.R. 1975. Weeds and domesticates: evolution in a man-made habitat. Economic Botany 29:99-107. Doggett, H. 1988. Sorghum. 2nd Edition. Tropical Agricultural Series. Longman Scientific, Essex. Eastin, J.D. and Lee, K. 1985. Sorghum bicolor. In: CRC Handbook of Flowering. IV. Halevy, A.H. (Ed.), pp. 367-375. CRC Press, Boca Raton, FL, USA. Eber, F., Chevre, A.M., Baranger, A., ValIee, P., Tanguy, X. and Renard. M. 1994. Spontaneous hybridization between a male-sterile oilseed rape and two weeds. Theoretical Applied Genetics 88:362-368. Ellstrand, N.C. 1992. Gene flow by pollen:implications for plant conservation genetics. Oikos 63:77-86. Ellstrand, N.C., and Hoffman, C.A. 1990. Hybridization as an avenue for the escape of engineered genes. Bioscience 40: 438-442. FAO. 1993. Production Yearbook. Food and Agriculture Organization of the United Nations. Rome, Italy. pp. 84-85. Futuyma, D.J. 1986. Evolutionary Biology. Sinauer, Sunderland. MA, USA. Gliddon, C. 1994. The impact of hybrids between genetically modified crop plants and their related species: biological models and theoretical perspectives. Molecular Ecology 3:41-44. Goodman, R.M. and Newell. N. 1985. Genetic engineering of plants for herbicide resistance: status and prospects. In: Engineered Organisms in the Environment: Scientific Issues. Halvorson. H.O., Pramer, D., and Rogul, M., (Eds.), pp. 47-53. American Society for Microbiology, Washington, D.C., USA. Holm, L.G., Plucknett, D.L., Pancho, J.V. and Herberger. J.P. 1977. The World's Worst Weeds. University Press of Hawaii, Honolulu.HI, USA. Jorgensen, R.N. and Andersen, B. 1994. Spontaneous hybridization between oilseed rape Brassica napus and weedy B. campestris (Brassicaceae): a risk of growing genetically modified oilseed rape. American Journal of Botany 81:1620-1626. Kigel, J. and Rubin, B. 1985. Sorghum halepense.In: CRC Handbook of Flowering. IV. Halevy, A.H. (Ed.), pp. 376-379. CRC Press, Boca Raton, FL, USA. Kirkpatrick, K.J. and Wilson, H.D. 1988. Interspecific gene flow in Cucurbita: C texana vs. C. pepo. American Journal of Botany 75: 519-527. Klinger, T., ArrioIa, P.E. and Ellstrand. N.C. 1992. Crop-weed hybridization in radish (Raphanus sativus L.): effects of distance and population size. American Journal of Botany 79:1431-145. Klinger, T., Elam. D.R. and Ellstrand, N.C. 1991. Radish as a model system for the study of engineered gene escape rates via crop-weed mating. Conservation Biology 5:531-535. Klinger. T. and Elistrand, N.C. 1994. Engineered genes in wild populations: fitness of weedcrop hybrids ofRaphanus sativus. Ecological Applications 4:117-120. Ladizinsky, G. 1992. Crossability relations. In: Distant Hybridization in Crops. Kalloo, G. and Chowdhury, J.B. (Eds.), pp. 15-31. Springer-Verlag, Berlin, Germany. Langevin, S.A., Clay, K. and Grace, J. 1990. The incidence and effects of hybridization between cultivated rice and its related weed red rice (Orvza sativa L.). Evolution 44: 1000-1008. National Research Council. 1989. Field testing genetically modified organisms: framework for decisions. National Academy Press, Washington, D.C., USA. Prescott-Allen, R. and Prescott-Allen, C. 1983. Genes From the Wild. International Institute for Environmental Development. Russell Press, Nottingham, UK. Raybould, A.F. and Gray, A.J. 1994. Will hybrids of genetically modified crops invade natural communities? Trends in Ecology and Evolution 9:85-89. Santoni, S. and Berville, A. 1992. Evidence for gene exchanges between sugar beet (Beta vulgaris L.) and wild beets: consequences for transgenic sugar beets. Plant Molecular Biology 20:578-580. Slatkin. M. 1987. Gene flow and the geographic structure of natural populations. Science 236, 787-792. Small, E. 1984. Hybridization in the domesticated- weed-wildcomplex. pp. 195-219. In: Plant Biosystematics. Grant, W.F. (Ed.). Academic Press, Toronto, Canada. Smith, M. and Adams, W.T. 1983. Measuring pollen contamination in clonal seed orchards with the aid of genetic markers. In: Proceedings 17th Southern Forest Tree Improvement Conference, University of Georgia, Athens, GA, USA, pp. 69-77. Till-Bottraud, I., Reboud, X., Brabant, P., Lefranc, M., Rherissi, B., Vedel, F.and Darmency, H. 1992. Outcrossing and hybridization in wild and cultivated foxtail millets: consequences for the release oftransgenic crops. Theoretical Application of Genetics 83:940-946. Umbeck, P.F. Barton, K.A., Nordheim, E.V., McCarty, J.C. Parrott, W.L. and Jenkins, J.N. 1991. Degree of pollen dispersal by insects from a field test of genetically engineered cotton. Journal of Economic Entomology 84: 1943-1950. Warwick, S. I. and Black, L.D. 1983. The biology of Canadian weeds. 61. Sorghum halepense (L.) Pers. Canadian Journal of Plant Science 63:997-1014. Wilson, H. and Manhart, J. 1994. Crop/weed gene flow: Chenopodium quinona Willd. and C. berlandieri Moq. Theoretical Application of Genetics 86:642-648. Copyright 1995 African Crop Science Society
The following images related to this document are available:Line drawing images[cs95021a.gif] [cs95021c.gif] [cs95021b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}