 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Some implications of genotype X environment interactions on sowing seed quality of sorghum A. TENKOUANO and P. SEREME Institut d'Etudes et de Recherches Agricoles, 03 BP 7192 Ouagadougou 03, Burkina Faso (Received 29 August 1995, accepted 23 January. 1996)
Code Number: CS96035
Sizes of Files:
Text: 29K
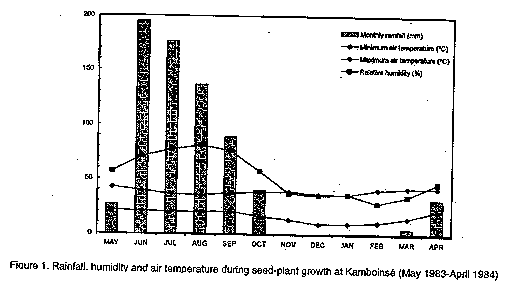
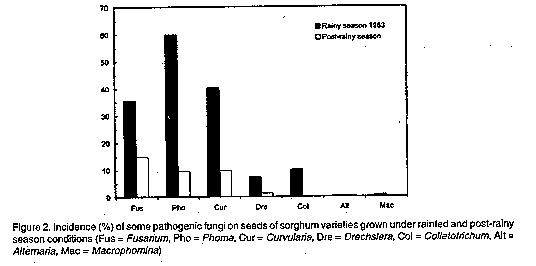
Graphics: Line Drawings (gif) - 9.4KABSTRACT Poor seed quality is a major constraint to the adoption of improved sorghum cultivars in West Africa because it results in poor stands. While genetic enhancement of this trait should not be overlooked, more immediate improvement of seed quality could result from better exploitation of the production environment. Under this postulate, seeds of thirty sorghum cultivars grown under normal rainfed (RS) conditions and under irrigation during the post-rainy (PR)season were compared for seed quality and seedling emergence and growth (grain quality affects products). Season effects accounted for a greater proportion of variance in phenotypic expression of cultivars than did genetic effects. Seeds from the PR harvest were less weathered and displayed greater grain weight, density and seedling emergence percentage compared to RS seeds. Furthermore, PR seeds produced more vigorous and faster growing seedlings than RS seeds. This work indicates that the post-rainy season, where possible, could be used to produce high quality seeds at reduced cost, since only supplemental irrigation is needed. Seed production in the post-rainy season would offer the added advantage of assuring isolation and reducing the risks of uncontrolled pollination. Key Words: Sorghum bicolor, seed quality, genetic effects, environmental effects RESUME La qualite des semences est une contrainte majeure a l'adoption des cultivars ameliors de sorgho en Afrique del'Ouest. Bien que l'amelior genetique de ce caractere soit envisageable, des resultats plus immediats pour plus immediats pourraient etre obtenus par une meilleure exploitation du milieu de production. Avec ce postulat, les recoltes d'hivernage et de contre saison de trente cultivars de sorgho ont fait l'objet d'une comparaison pour leur qualite grainiere et leur aptitude semenciere. L'etude revele la preponderance des sources de semences sur les effets genetiques dans la variation de l'expression phenotypique des cultivars. Les semences de contre-saison sont moins deteriorees et ont un mellieur poids, une meilleure densite et une meilleure levee que les semences d'hivernage. Les plantules provenant des semences de contre-saison sont aussi plus vigoureuses que celles produites par les semences d'hivernage. Ce travail montre que l'on devrait exploiter la periode post-hivernale pour produire des semences de bonne qualite a moindres frais puisque que seule une irrigation d'appoint est requise. La production semenciere encontre-saison permettrait aussi d'assurer un bon isolement et de reduire les risques de pollinisation incontrolee. Mots Cles: Sorghum bicolor, qualite semenciere, effets genetiques, effets environnementaux INTRODUCTION Poor crop stands are often cited as a major constraint to the production of rainfed cereals in many arid and semi-arid areas in the world (Stoop et al., 1982). Suboptimal crop establishment results from the additive or interactive effects of several seed or nonseed factors. For example, the morphological, histological and physiological characteristics of the seed determine its intrinsic ability to germinate and produce a seedling that will emerge through the soil surface and exhibit fast and vigorous growth. While sowing seed quality could be enhanced through genetic manipulation, more immediate improvement could be obtained through better exploitation of the environment in which the seed plant is grown. Thus, crop management practices, such as seedbed preparation and sowing time and method, determine the physical and chemical environment for seed germination and seedling emergence and growth. Thus, suboptimal seed development or failure to ensure adequate soil conditions will result in failure of seed germination, failure of seedling emergence and low survival of emerged seedlings (Soman et al., 1984). The relationships of seedling emergence, survival, and growth to sowing seed quality is known for most economically important crops in the industrialised world where seed production and priming is an essential and distinct component of crop production. However, in non-industrial regions of the world, e.g. the Sahel of West Africa, farmers do not differentiate between seed and grain as the grain from the current crop is saved as seed for the next crop. In those traditional production systems, farmers routinely resort to several sowings before acceptable plant stands can be achieved (Catherinet et al., 1973). The high cost of modern seed production is currently prohibitive for non-industrial countries because of a lack of a seed market in those regions. Therefore, alternative methods of producing high quality seeds at reduced cost must be sought. In West Africa, sorghum (Sorghum bicolor [L] Moench) is cultivated during the normal season of crop growth under rainfed conditions. In some areas, however, it is possible to grow an additional crop under supplemental irrigation during the period following the rainy season (post-rainy season), before photoperiod becomes a constraint. In these conditions, seeds harvested from PR nurseries were observed to produce more vigorous seedlings than seeds from the RS crop although the basis for such differences were not investigated (ICRISAT, 1982). Also, whether genetic variation exists among sorghum cultivars for this differential seed performance is not known. The objective of this study, therefore, was to examine the effects of interaction between seed plant genotype and cropping environment on seed quality and seedling growth characteristics of improved and farmers' varieties of sorghum. MATERIALS AND METHODS Twenty photoperiod-insensitive breeding lines of sorghum from the International Crop Research Institute for the Semi-Arid Tropics (ICRISAT) and ten photoperiod-sensitive indigenous cultivars were used in the study. The ICRISAT lines were derived from exotic (Caudatum) x local (Guineense) or exotic x exotic crosses. Seven of the breeding lines were early-maturing (<70 days to anthesis), 7 were intermediate (70-80 days) and 6 were late-maturing (> 80 days). Indigenous cultivars were derived from mass selection of farmers' varieties and included 5 early-maturing cultivars from the Sahelian agroecological zone and 5 late-maturing cultivars from the Sudanjan agroecological zone of West Africa. These materials were grown at Kamboinse Agricultural Research Station (13'N, 2'W), in Burkina Faso. Planting was done at approximately 15 day intervals for late, intermediate and early maturing cultivars to meet maturity requirements. Preplanting fertilisation (15N:23 P205:15 K20) was done at the rate of 100 kg ha^-1 and urea (46% N) was applied as side-dressing in two split-doses of 50 kg ha^-1 each at 30 and 60 days after planting. Rainfall received at Kamboinse was adequately distributed (Fig. 1) and supplemental irrigation was provided only for the post-rainy season crop as needed. Harvest was done in October 1983 and March 1984 for the RS and PR crops, respectively. Seeds from these plots were evaluated for grain weathering, endosperm texture, and 100 kernel weight using methods previously described (Rao and Williams, 1980; House, 1980). The blotter paper method developed by the International Seed Testing Association (ISTA, 1966) was also used to assess the extent of seed colonisation by specific pathogenic fungi. One hundred kernels were immersed in 100 ml of water in a graduated cylinder, and grain density was obtained as the ratio of 100 seed weight to the volume of water displaced by the 100 kernels. The kernels were then disposed between layers of moist Whatman paper at room temperature for 48 hours after which a count was made to determine percent germination. Thirty six kernels were sown in the greenhouse, in aluminum pots filled with a mixture of sand and farm manure in a 3:1 ratio. Percent emergence was assessed 7 days after sowing (DAS) and divided by germination percentage to obtain adjusted seedling emergence percentage. Seven and 14 DAS, the seedlings were cut at the soil line and the roots were washed to remove soil particles. Lengths of both aerial and underground seedling parts were measured. These fractions were then ovendried at 70 C for 72 hours to determine their dry weight. Data were recorded from six replications of a split-plot arranged within a randomised block design. Main plot and subplot treatments consisted of the cultivars and seed sources, respectively. RESULTS Main and interaction effects were significant for all traits, except grain density (Tables 1 & 2). Interseasons effects accounted for considerably greater variance than any other source of variation, thereby indicating the preponderance of seed plant environment in the determination of seed quality factors. A larger proportion of the variance due to either cultivar or interaction effects resulted from differences between rather than within groups of cultivars, which also stresses that seed quality may be more related to seed plant environment than to plant genotype. The data further indicate that seeds obtained from plants grown during the post-rainy season had higher quality than those produced during the rainy season, regardless of plant genotype (Tables 3&4, Fig.2). Also, significant correlations were found between seed quality and seedling emergence and growth (Table 4). DISCUSSION The sorghum panicle and grain is often subject to attack by a complex of mycoflora which induce lesions that alter seed structure and composition hence seed quality (Castor and Figure 1. Rainfall, humidity and air temperature during seed-plant growth at Kamboinse (May 1983-April 1984) Figure 2. Incidence (%) of some pathogenic fungi on seeds of sorghum varieties grown under rainfed and post-rainy season conditions (Fus = Fusarium, Pho = Phoma, Cur = Curvularia, Dre = Drechslera, Col = Colletotrichum, Alt = Alternaria, Mac = Macrophomina) Frederiksen, 1980; Louvel, 1983a&b; Rao and Williams, 1980). TABLE 1. Mean squares from the analysis of variance for seed quality characteristics of sorghum varieties grown during rainy and post-rainy season conditions
Source of variation
Df Grain Endosperm 100
weath- texture kernel Grain Seedling
weight density emergence
------------------------------------------------------------
Replications 5 1.24 1.64 O.10 0.33 1733.94
Cultivars 29 2.01** 2.76** 2.24** 0.06 1400.13**
Inter-group 4 2.43** 2.24** 4.69** 0.03 4056.37**
Intra-group 25 1.94** 2.84** 1.85** 0.07 975.13**
Error A 145 0.30 0.29 0.10 0.05 302.59
Seasons 1 87.03** 26.14** 44.07** 1.13** 3659.45**
Cultivars x Seasons
29 1.27** 0.55** 0.65** 0.07** 1017.18**
Inter-group 4 1.31** 0.66** 0.61** 0.10** 2575.64**
Intra-group 25 1.26** 0.53** 0.66** 0.07** 767.83**
Error B 150 0.28 0.18 0.06 0.04 214.17
**: F-test significant at the 0.01 probability level.TABLE 2. Mean squares from the analysis of variance for seedling growth characteristics of sorghum varieties grown during rainy and post-rainy season conditions
Source of variation
Df Seedling length Seedling dry weight
7 DAS 14 DAS 7 DAS 14 DAS
--------------------------------------------------------------
Replications 5 6309.72 21623.52 306.38 6454.78
Cultivars 29 1285.45* 6950.12" 35.39* 1055.91*
Inter-group 4 3536.82'* 6776.69 70.01** 2139.74**
Intra-group 25 925.23 6977.87** 29.85 882.49
Error A 145 839.95 3706.94 20.42 655.20
Seasons 1 51816.00** 195673.47** 1173.61** 36100.07**
Cultivars x Seasons
29 1894.09** 6996.07** 23.73 967.72**
Inter-group 4 3887.41 15331.06** 41.59* 1822.48*
Intra-group 25 1575.16 5665.67** 20.87 830.96
Error B 150 709.64 2888.09 16.15 564.55
-*, **: F-test significant at the 0.05 and 0.01 probability levels, respectively. Invasion of the panicle occurs as early as anthesis and is favoured by hot and humid weather conditions (Castor and Frederiksen, 1980; Denis and Girard, 1980) such as those which prevailed from mid July to mid September at Kamboinse (Fig.1). Conversely, relatively cooler and drier weather conditions were recorded during the post-rainy season at Kamboinse (Fig. 1), as is often the case in the Sahel (ICRISAT, 1984). Also, increased air velocity and turbidity normally occur within crop populations during the post-rainy season in the Sahel due to the Sahara desert-blown Harmattan (ICRISAT, 1984). This prevents stabilisation of thermal and hygrometric regimes, which, in turn, does not allow for conditions that are favourable for fungal development. These circumstances account for the reduced fungal ingress of sorghum kernels for PR-grown sorghum compared to their RS counterparts (Table 3). Variability in sorghum resistance to grain weathering has often been equated with variability m maturity cycle. In this study, resistant and susceptible reactions were recorded in all maturity groups (Table 3). Results were not supportive of the commonly accepted association of susceptibility with earliness. This was attributed to the fact that actual maturation conditions rather than length of growth cycle were the determinant factors of fungal ingress in developing kernels (Denis and Girard, 1980). In fact, our results failed to identify any phenological attribute that could be linked to resistance to weathering. Glueck and Rooney (1980) further determined that resistance to weathering may be more related to the physiological and chemical properties of the kernel such as water movement and presence or absence of certain phenolic compounds in grain pericarp. This is of major and practical significance since the possibility of developing early maturing yet grain mould resistant varieties is suggested. Endosperm texture is determined by genotype, and grain fill, a physiological process driven by translocation of photosynthates to the grain (Newton et al., 1983) coupled with seed moisture elimination from the grain through transpiration. Since sorghum cultivars differ in photosynthetic capacity and translocation efficiency (Muchow and Wilson, 1976), cultivar differences for endosperm texture found in this study (Table 3) and in others (Rooney and Miller, 1982) were expected. TABLE 3. Seed quality characteristics of sorghum varieties grown under normal rainfed (RS) and irrigated post-rainy (PR) season conditions
Character
------------------
Seed Maturity and race classification
source ------------------------------------------------------
Early Medium Late Early Late
Caudatum Caudatum Caudatum Guinea Guinea
-------------------------------------------------------------
Grain weathering^a
RS 2.7+/-0.33 2.4+/-0.61 2.2+/-0.62 2.4+/-0.60 2.0+/-0.74
PR 1.3+/-0.24 1.5+/-0.43 1.5+/-0.37 1.1+/-0.12 1.1+/-0.20
Endosperm texture^b
RS 2.7+/-0.29 2.6+/-0.42 2.6+/-0.45 2.0+/-0.28 3.4+/-0.42
PR 2.2+/-0.27 2.1+/-0.19 2.1+/-0.34 1.7+/-0.29 2.4+/-0.79
100 kernel weight(g)
RS 2.0+/-0.15 2.2+/-0.12 1.9+/-0.23 2.2+/-0.29 2.6+/-0.63
PR 2.8+/-0.45 3.0+/-0.28 2.4+/-0.31 2.6+/-0.56 3.6+/-0.50
Grain density (g.cm^-3)
RS 1.0+/-0.06 1.0+/-0.07 1.1+/-0.14 1.2+/-0.06 1.0+/-0.04
PR 1.2+/-0.04 1.2+/-0.06 1.2+/-0.10 1.1+/-0.07 1.2+/-0.13
Seedling emergence (%)
RS 29.3+/-7.44 47.7+/-11.34 51.3+/-10.06 71.0+/-3.35 48.0+/-
4.23
PR 71.1+/-7.52 69.7+/-9.87 56.3+/-11.41 77.6+/-6.02 75.8+/-
6.37
^a: On a 1 to 5 scale where 1 = healthy and 5 = severely weathered Because higher transpiration rates are associated with increased air temperatures and reduced air humidity (ICRISAT, 1984), it was anticipated that increased packing of translocated metabolites in the seeds would occur in the post-rainy season compared to the rainy season, hence the results obtained. Similarly, cultivars and season differences for grain weight were consistent with earlier reports (Liang et al. 1969; Singh and Baghel, 1977; Glueck and Rooney, 1980). Both cultivar and season effects were not significant for grain density in our study. This contrasted with the results of Glueck and Rooney (1980) which indicated that the test weight, seed density arid several other physical and chemical properties of sorghum cultivars were affected by the environment in which they were grown. Seedling emergence was generally better for local cultivars than for breeding lines, among which little difference was observed. Season effect for this trait was large and significant for virtually all early and medium maturing breeding lines and for one group of local cultivars. Interestingly, these local cultivars were originally collected from some of the drier parts of the Sahel and, therefore, have reduced growth period compared to the other locals. These materials were also similar to the short- and medium- cycle breeding lines for grain vitreosity, since they produced starchy seeds under rainfed conditions. No significant cultivar or source effect was found root growth characteristics (data not shown). In contrast, both main and interaction effects were significant for the length and the dry weight of seedling aerial parts both 7 and l 4 DAS (Table 2). Seedlings produced from PR seeds elongated faster and accumulated more dry matter than their RS counterparts particularly for early and medium maturing lines. However, the magnitude of within cultivar season differences was markedly reduced 14 DAS (Table 4). Thus seed source and cultivar effects appeared to be critical for initial seedling growth. These factors seem to have little influence on the growth of seedlings after they have developed sufficient root systems and no longer depend on the reserves accumulated in the endosperm during seed maturation. Seed characteristics have sometimes been used as predictors of seedling establishment since it has been suggested that poor grain fill and sanitary status reduce seedling growth (Glueck and Rooney, 1980; Louvel, 1983 a&b). TABLE 4. Seedling growth characteristics of sorghum varieties produced under normal rainfed (RS) and irrigated post-rainy (PR) season conditions
Character
Seed Maturity and race classification
source
Early Medium Late
Caudatum Caudatum Caudatum
-----------------------------------------------------
Seedling length (mm)
7 DAS^a RS 56.3+/-17.74 73.6+/-15.24 72.7+/-10.72
PR 99.0+/-11.06 105.6+/-18.14 90.5+/-10.32
14 DAS RS 228.7+/-51.61 267.0+/-23.10 253.5+/-23.66
PR 318.7+/-19.40 308.6+/-33.68 300.8+/-18.32
Seedling dry weight (mg)
7 DAS RS 8.4+/-1.59 8.0+/-2.39 7.5+/-1.26
PR 12.6+/-1.76 13.7+/-1.91 10.8+/-1.95
14 DAS RS 46.6+/-16.02 56.9+/-10.47 49.8+/-8.71
PR 79.6+/-10.07 76.0+/-11.74 68.2+/-12.55
Table 4 contd./
Character
Seed Maturity and race classification
source
Late Early Late
Caudatum Guinea Guinea
--------------------------------------------------------------
Seedling length (mm)
7 DAS^a RS 72.7+/-10.72 84.0+/-8.12 88.8+/-13.48
PR 90.5+/-10.32 91.6+/-12.78 100.0+/-7.04
14 DAS RS 253.5+/-23.66 283.4+/-29.45 280.6+/-20.66
PR 300.8+/-18.32 284.2+/-16.34 319.4+/-29.99
Seedling dry weight (mg)
7 DAS RS 7.5+/-1.26 8.8+/-1.33 11.6+/-0.80
PR 10.8+/-1.95 10.4+/-0.80 13.6+/-2.06
14 DAS RS 49.8+/-8.71 59.0+/-8.65 64.2+/-5.53
PR 68.2+/-12.55 62.4+/-7.17 83.0+/-7.69^a: DAS - days after sowing TABLE 5. Correlation between seed quality indicators and seedling emergence and growth
Seed characteristic
Seedling
emergence Seedling length Seedling dry
weight
------------------------------------------ Grain
weathering 0.38** 7 DAS 14 DAS 7 DAS 14 DAS
Endosperm texture 0.35** 0.30** 0.34** 0.27** 0.27**
100 kernel weight 0.23** 0.18** 0.24** 0.06 0.24**
Grain density 0.19** 0.17** 0.25** 0.31** 0.32**
0 08 0.07 0.03 0.08 **: Significant at the 0.01 probability level However, the correlation coefficients calculated in this study were quite small (Table 5), thus limiting the predictive value of single seed trait analysis on seedling development. A commonly held view is that the photoperiod requirements of indigenous sorghum materials assure that they mature under conditions that are not favourable to disease and pest damage. This study shows that although these materials were less affected by weathering under rainfed conditions, they did not perform much better than exotic materials of comparable growth cycle because of their failure to fill their grain adequately (Table 3). Thus, photoperiod sensitivity does not guarantee that the plant will produce seeds of acceptable quality. With supplemental irrigation in the post-rainy season, virtually no differences were found in the mean performance of the various groups of material s, although seedling emergence of late maturing exotics remained lower than that of the other groups. CONCLUSION This study confirms that seed production environment influences seed performance but restricts that influence to the early events of emergence and growth. It is this early growth and survival that ultimately determines crop stand. Our results suggest that the post-rainy season, where possible, could be used to produce high quality seeds, particularly for improved cultivars. Seed production in these conditions will be cost effective, since only supplemental irrigation will be required. Furthermore, seed production in the post-rainy season would assure isolation and reduce the risks of uncontrolled pollination. The preponderance of environmental causes of variation over genetic effects does not suggest that the importance of genotype to the quality of sowing seeds should be minimised. This work suggests that the genetic make-up of a cultivar controls the sowing quality of the seeds it produces. However, the relationships of genetics to the sowing quality of sorghum kernels may not be the same as that of genetics to end-use quality of the kernel (Rooney and Miller, 1982). However, breeding for end-use quality of seed has often resulted in modification of sowing seed quality, which shows that direct or indirect selection for sowing seed quality is also feasible. ACKNOWLEDGEMENTS The authors thank Dr. C.M. Pattanayak, former Team Leader, International Crop Research Institute for the Semi-Arid Tropics (ICRISAT), Burkina Faso, for kindly providing guidance and partial financial support for this research. We are also indebted to Drs. L.W. Rooney. L.E. Clark, and F.R. Miller, Texas A&M University/Texas Agricultural Experiment Station, U.S.A., for their comments and suggestions during the preparation of the manuscript. REFERENCES Castor, L.L. and Frederiksen, R.A. 1980. Fusarium and Curvularia grain mold in Texas. In: Sorghum Diseases: A World Review. Proceedings of the International Workshop on Sorghum Diseases, 11-15 Dec. 1978, Hyderabad. Williams, R.J. and Frederiksen, R.A. (Eds.), pp. 93-102. ICRISAT, Patancheru, India. Catherinet, M.D., Dumont, S. and Mayaki, A.A. 1973. Le mil et le sorgho dans l'agriculture du Niger. A. Besoins, sol et climat, facteurs limitants. L'Agronomie Tropicale 18: 108113. Denis, J.C. and Girard, J.C. 1980. Factors affecting the development of sorghum grain molds in Senegal. In: Sorghum Diseases: A World Review. Proceedings of the International Workshop on Sorghum Diseases, 11 - 15 Dec. 1978, Hyderabad. Williams, R.J. and Frederiksen, R.A. (Eds.), pp. 144-153. ICRISAT, Patancheru, India. Glueck, J.A. and Rooney, L.W. 1980. Chemistry and structure of grain in relation to mold resistance. In: Sorghum Diseases: A World Review. Proceedings of the International Workshop on Sorghum Diseases, 11 - 15 Dec 1978, Hyderabad. Williams, R.J. and Frederiksen, R.A. (Eds.), pp. 119-140. ICRISAT, Patancheru, India. House, L .R. 1980. A Guide to Sorghum Breeding. ICRISAT, Patancheru, India, 238 pp. ICRISAT, 1982. Annual Report. Sorghum Breeding. ICRISAT, Ouagadougou, Burkina Faso. 262 pp. ICRISAT, 1984. Agrometeorology of Sorghum and Millet in the Semi-Arid Tropics. Proceedings of the International Symposium, 15-20 Nov. 1982, ICRISAT Center India. Patancheru, India. 322 pp. ISTA, 1966. International rules for seed testing. Proceedings of the International Seed Testing Association 31: 1-152. Liang, G.H.L., Walter, T.L., Nickell, C.D. and Koh, Y.O. 1969. Heritability estimates and interrelationships among agronomic traits in grain sorghum, Sorghum bicolor (L.) Moench. Canadian Journal of Genetics and Cytology 11: 199-208. Louvel, D. 1983a. Phytopathologie du sorgho. Les moisissures des grains. Etude du pouvoir pathogene de quelques elements de la mycoflore. Coll. Etudes Techniques du CNRA, Dakar, Senegal. 40 pp. Louvel, D. 1983b. Phytopathologie du sorgho. Vigueur des plantules et mycoflore du grain. Revue bibliographique. Coil. Etudes Techniques du CNRA, Dakar, Senegal. Muchow, R.C. and Wilson, G.L. 1976. Photosynthetic and storage limitations to yield in Sorghum bicolor (L.) Moench. Australian Journal of Agricultural Research 27:489-500. Newton, R.J., Meckenstock, D.H. and Miller, F.R. 1983. Soluble carbohydrates in developing sorghum caryopses. Crop Science 23:80-82. Rao, K.N. and Williams, R.J. 1980. Screening for sorghum grain mold resistance at ICRISAT. In: Sorghum Diseases: A World Review. Proceedings of the International Workshop on Sorghum Diseases, 11-15 Dec 1978, Hyderabad. Williams, R.J. and Frederiksen, R.A. (Eds.), pp. 103-108. ICRISAT, Patancheru, India. Rooney, L.W. and Miller, F.R. 1982. Variation in the structure and kernel characteristics of sorghum. In: Proceedings of the International Symposium on Sorghum Grain Quality, 28-31 Oct 1981, ICRISAT Center, Patancheru, India. Rooney, L.W. and Murty, D.S. (Eds.), pp. 143-162. ICRISAT Patancheru, India. Singh, R.P. and Baghel, S.S. 1977. Yield components and their implication to selection in sorghum. Indian Journal of Genetics and Plant Breeding 37:62-67. Soman, P., Peacock, J.M. and Bidinger, F.R. 1984. A field technique to screen seedling emergence in pearl millet and sorghum through soil crust. Experimental Agriculture 20: 327334. Stoop, W.A., Pattanayak, C.M., Matlon, P.J. and Root, W.R. 1982. A strategy to raise the productivity of subsistence farming systems in the West African semi-arid tropics. In: Sorghum in the Eighties.- Proceedings of the International Symposium on Sorghum, 27 Nov. 1981, ICRISAT Center, Patancheru, India. House, L.R., Mughogho, L.K. and Peacock, J.M. (Eds.), pp. 519-526. ICRISAT Center, Patancheru, India. Copyright 1996 The African Crop Science Society The following images related to this document are available:Line drawing images[cs96035a.gif] [cs96035b.gif] |
| |||||||||

{kind=link}
{kind=link}