 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Cardinal temperatures and thermal times for vernalisation in carrot cv. "Nantes"
E.M. KAHANGI, J. A. CHWEYA^1 and L.S. AKUNDABWENI^1 Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P.O. Box 62000, Nairobi, Kenya. ^1 Department of Crop Science, University of Nairobi, P.O. Box 29053, Nairobi, Kenya. (Received 22 February 1995; accepted 30 October 1995)
Code Number: CS96040
Sizes of Files:
Text:20.4K
Graphics: Line Drawings (gif) - 8.8KABSTRACT
Effect of temperatures on bolting and flowering, measured as days to internode extension (bolting) and to flower bud visibility, respectively, were studied in carrot (Daucus carota) cultivar (cv.) "Nantes" grown in a controlled environment. All carrot plants raised from stecklings subjected to 10 weeks chilling at 3, 6 and 9 C bolted and flowered under subsequently warmer temperatures (18 C). However, only 33% of the stecklings given 1 and 12 C temperature pretreatments for a similar duration flowered. No flowering was observed for plants whose stecklings were pre-treated at 15 C for 10 weeks or those exposed to 1 C for 5 weeks. Plants chilled for 10 weeks required less days to bolting and flowering (P < 0.05) than those chilled for 5 weeks. Rates of bolting and flowering increased linearly with temperatures from 1 to 6 C and declined linearly above 6 to 12 C. The linear relationship enabled the optimum temperatures for vernalisation to be derived as 6 C for both bolting and flowering. Base temperatures of -2.9 and -1.7 C and maximum temperatures of 16.97 and 16.27 C for bolting and flowering, respectively, were also predicted. The minimum thermal time for vernalisation in degree Celcius days (Cd) for any bolting and flowering to occur in this cultivar of carrot was 126 Cd and the optimum for 100% bolting and flowering was about 400 Cd. Key Words: Bolting, Daucus carota, flowering, stecklings RESUME
L'effet de temperatures sur l'elongation des entre-noeuds et la floraison, mesure respectivement en nombre de jours d'elongation d'entrenoeuds et d'apparition de boutons floraux, etait etudie sur le cultivar de carotte "Nantes" plantes en milieu controle. Tousles plants de carotte exposes a 10 semaines de refroidissement de 3, 6, et 9 C se sont etioles et ont fleuri a des temperatures subsequentes chaudes (18 C). Cependant, seulement 33 % de plants ayant recus des pretraitements de temperatures de 1 et 12 C pendant une periode similaire. Les plantes qui n'ont pas fleuri sont ceux qui ont renu un pretraitement de 1 C pendant 10 semaine ou ceux qui etaient exposes a la temperature de 1 C pendant 5 semaines. Les plants refroidis pendant 10 semaines exigeaient peu de jours pour l'elongation des entrenoeuds et la floraison (P<0.05) par rapport a ceux refroidis pendant 5 semaines. Les taux d'elongation et de floraison augmentaient lineairement a des temperatures de 1 a 6 C, et decroissaient lineairement au dessus de 6 a 12 C. La relation lineaire a permi d'ajuster l'optimum de temperatures de vernalisation a 6 C pour aussi bien l'elogation d'entrenoeuds que la floraison. De meme, il etait bien possible de predire que pour l'elongation et la floraison, les temperatures extreme etaient -2.9 et 16.97, et -1.7 et 16.27 C respectivement. Le temps thermal minimum en degres celcius parjour de vernalisation pour que l'elongation. et la floraisonaient lieu sur ce cultivar de carotte etait de 126 Cd tandis que l'optimum pour avoir 100% d'elongation et de floraison etait environ 400 Cd. Mots Cles: Elongation d' entrenoeuds, Daucus carotta, floraison, stecklings INTRODUCTION It has long been known that the stimulus for bolting and floral initiation in carrot (Daucus carota L.) is exposure to low temperatures (Gassner, 1918), a phenomenon referred to as vernalization. High percentage of flowering is the basis for economic seed yield and is, therefore, of great importance to breeders and seed producers. Previous studies under controlled environments have shown that different carrot cultivars have a qualitative vernalisation response to temperatures between 0 and 10 C and that lengthening the exposure from two to eight weeks increased the response (Sakr and Thompson, 1942; Dickson and Peterson, 1958). However, differences in the effectiveness of these temperatures have been reported only in cv. "Chantenay Red Cored" (Atherton et al., 1990). In this cultivar the base, optimum and maximum temperatures for vernalisation were estimated to be -1", 6.5 and 16.0 C, respectively. In the temperate zone, carrot plants grown for seed production are exposed to natural chilling temperatures during winter seasons to subsequently flower in spring. In contrast, chilling temperatures are lacking in the tropical lowlands. Even in the tropical highlands, it is not clearly understood whether the highlands provide sufficient chilling for satisfactory flowering in carrots for the purpose of seed production. Before the confirmation of this, the effectiveness of different temperatures and their durations on vernalisation in carrot should be established. This study, therefore, evaluated the vernalisation response in carrot cv. "Nantes", the most popularly grown cultivar in Kenya. MATERIALS AND METHODS Carrot roots (stecklings) were raised at the Jomo Kenyatta University of Agriculture and Technology (JKUAT) farm using the recommended agronomic methods. Imported carrot seeds of cv. "Nantes" were sown in January, 1991, the stecklings harvested three months later, and the best roots for the experiment selected. These were prepared for cold storage by washing off excess soil, trimming their leaves to 2.5 cm above the crown and dipping them in a fungicide solution containing Captan (3g l^-1 of water) and Ridomil (2g l^-1 of water) for 15 minutes to kill any soil borne fungal diseases. The stecklings were finally placed in perforated plastic bags with sterilised sawdust to prevent desiccation during cold storage. Temperature treatments were 1 C, 3, 6, 9, 12 and 15 C for 5 and 10 weeks durations. These treatments were selected basing on one year preliminary work. The roots were placed in incubators set at respective temperatures. Upon completion of temperature treatments, 24 stecklings for each vernalisation temperature and duration were potted in 15 cm diameter plastic pots containing sterilised forest soil and sand at the ratio of 2:1. The potted roots with temperature treatments were then arranged in a completely randomised design in daylight growth chambers (Growth Master Obata Manufacturing Co. LTD., Model RGM- 1500RH) set at 18 C. Each treatment was replicated four times. After sprouting, the plants were supplied with 2g diammonium phosphate fertilizer per pot. Days taken from planting to flowerstalk visibility (bolting) and to flowering (just before the main umbel opened and the umbellets had just started to show creamish petals inside^) were recorded for each plant in every treatment. Number of days to the two events and the proportion of plants that bolted and flowered were worked out after a six months period of observation. RESULTS Bolting and flowering response to different temperature pre-treatments. One hundred percent of the plants treated for 10 weeks at 3, 6 and 9 C subsequently bolted and flowered in daylight growth chamber set at 18 C (Table 1 ). But in those plants that were given I anti 12 C temperature pre-treatments for a similar duration, only 33% flowered. By the end of the observation period (6 months) no bolting or flowering occurred in any plant that had been exposed to temperatures of 15 C for 5 or 10 weeks or those exposed to C for 5 weeks (Table 1 ).
The shortest time to bolting was observed in plants
vernalised at 6 C for 10 weeks and the longest time was for
the 12 C for 5 weeks treatment (Table 1). Plants chilled for
10 weeks took significantly (P
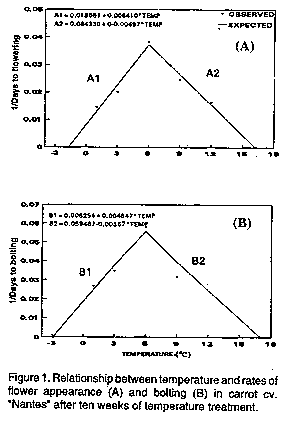
Cardinal temperatures for bolting and flowering. The
reciprocals of the mean days taken from the end of 10 weeks
temperature treatment to bolting and flower appearance at the
18 "C growth chamber (Table 1) were taken as measurements of
the rate of progress to each of the two events. The rates so
obtained were regressed on temperature using the method of
Atherton et al. (1990). Linear relationships emerged
when separate regressions were calculated for the sub- and
above optimum (supraoptimum) temperature ranges (Fig. 1).
Rates increased with temperatures between 1 and 6 C and
declined with temperatures between 6 and 12 C. Extrapolation
of the lines (Fig. 1) showed that carrot cv. "Nantes" has an
optimum vernalisation temperature (T[o]) of about 6 C for both
bolting and flowering. A base temperature (T[b]) and maximum
(T[m]) of about -2.9 and 17.0 C for bolting, respectively, and
-1.7 and 16.2 C for flowering were predicted. Cardinal
temperatures were calculated from the 10 weeks (70 days)
chilling durations, where more plants flowered, than the 5
weeks duration.
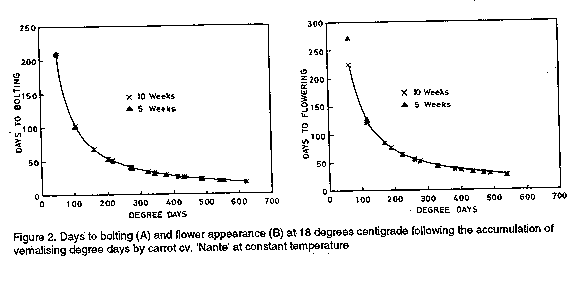
Thermal time for bolting and flowering. Thermal times
for vernalisation was calculated using Craigon et al.'s
(1990) method. The vernalising degree celcius days (Cd)
were worked out for each fixed temperature in the sub-optimum
and supra-optimum range using 70 days data (Table 1). When
vernalising degree days thus obtained were plotted against
days to bolting and flowering Craigon et al. (1990)
"model curves" (Fig. 2) were established.
The validity of the model curves derived from the 70 days
data for predicting flowering and bolting responses to other
temperature durations was tested using the 5 weeks (35 days)
data (Table 1).
TABLE 1. Effects of different temperatures, and
temperature duration treatments, on subsequent bolting
and flowering in carrot cv 'Nantes'
The time taken from the end of vernalisation to either bolting or
flowering decreased with increasing vernalisation times (Fig. 2). The
highest rate of bolting and flowering was between 75 and 250 Cd (.Fig. 2).
Acceleration of bolting and flowering continued as thermal time of
vernalisation increased beyond 250 Cd, but was less marked (Fig. 2).
Thermal times calculated from these relationships showed that carrot cv.
"Nantes" required a thermal time of vernalisation of about 400 Cd for 100%
flowering to occur.
DISCUSSION
Bolting and flowering in carrot cv. "Nantes" occurred at a common optimum
temperature of about 6 +/- 1 C. These observations were similar to those
made in carrot cv. Chantenay Red Cored" in which the optimum temperature
for the induction of bolting and flowering coincided at about 6.5 C
(Atherton et al., 1990). The extrapolated minimum temperatures for
bolting and flowering (-2.9 and -1.7 C, respectively) in cv. "Nantes" were
slightly different and so were the extrapolated maximum temperatures for
the two events (17.0 and 16.2 C for bolting and flowering, respectively).
Bolting and flowering in carrot have been reported to be independent
physiological processes (Hiller et al., 1979) and this could be the
reason why each of the two events had different minimum and maximum
temperatures.
The optimum and minimum temperatures obtained in this study fall within
the range of effective temperatures for vernalisation reported for most
pant species*(Chouard, 1960; Gallagher, 1979). On the other hand, the
maximum temperatures for vernalisation in carrot observed in this study are
slightly higher than those reported for most plant species (12 and 14 C)
(Chouard, 1960; Gallagher, 1979) but are in agreement with maximum
vernalisation temperatures observed in other cultivars of carrot ( Hiller
and Kelly, 1979; Atherton et al., 1990).
Earlier studies showed that carrots have a qualitative vernalisation
response to temperatures between 0 and 10 C (Sakr and Thompson, 1942;
Dickson and Peterson, 1958; Hiller and Kelly, 1979). But the reports did
not reflect on the differences in the effectiveness of these temperatures.
They, however, confirmed that lengthening the exposure from two to ten
weeks increases vernalisation response. Atherton et al. (1990) were
the first to report on effectiveness of different vernalisation
temperatures in carrot cv. "Chantenay Red Cored". The effectiveness of the
temperatures applied for both carrot cultivars increased from base to
optimum and decreased from optimum to maximum temperatures for
vernalisation.
Rate of vernalisation and temperatures showed linear relationships both
below and above the optimum. Such relationships are pre-requisite to the
use of thermal time to predict plant responses to the environment
(Garcia-Huidobro et al., 1982).
A minimum thermal time for vernalisation of about 135 Cd was observed
in this experiment. This induced about 17% of those individuals in the
population with the lowest vernalisation requirement to bolt and flower.
Atherton et al. (1990) observed a minimum thermal time of about 126
Cd in carrot cv. "Chantenay Red Cored". On the other hand, a thermal time
of vernalisation of about 400 Cd was required to obtain 100% bolting and
flowering in cv. "Nantes" while Atherton et al. (1990) reported
about 350 Cd in cv. "Chantenay Red Cored".
Model curves were established that predicted the same flowering and
bolting responses of mature carrot stecklings vernalised at different
durations (5 and 10 weeks). Consequently, a unit increase in temperature
anywhere within the sub-optimum range of vernalising temperatures was
equally effective in increasing the rate of progress towards flowering and
bolting. Conversion of temperatures for the plants vernalised at
supra-optimum temperatures to their sub-optimum effective temperatures
following Craigon et al. (1990) method, also predicted closely the
times of flowering and bolting. This agrees with Craigon et al.'s (1990)
findings in "Chantenay Red Cored" carrot and meets the basic criteria
or applying the thermal time procedure of Garcia-Huidobro et al.
(1982) to environments with varying temperatures. These findings will
be useful in predicting from meteorological records whether the Kenya
highlands have adequate chilling for vernalisation in carrot.
ACKNOWLEDGEMENTS
This work was supported by a grant from DAAD and this is gratefully
acknowledged.
REFERENCES
Arnold, C.Y. 1960. The determination and significance of the base
temperature in linear heat unit system. Proceedings of the American
Society for Horticultural Science 74:430-445.
Atherton, J.G., Craigon, J. and Basher, E., 1990. Flowering and bolting in
carrot. Juvenility, cardinal temperatures and therm, times for
vernalization. Journal of Horticultural Science 65:423-429.
Chouard, P. 1960. Vernalization and its relation to dormancy. Annual
Review of Plant Physiology I 1:191-238.
Craigon, J., Atherton, J.G. and Basher, E.A. 1990. Flowering and bolting in
carrot. II. Prediction in growth rooms, glasshouse and field environments.
Journal of Horticultural Science 65:547-554.
Dickson, M.H. and Peterson, C.E. 1958. Hastening greenhouse seed production
for carrot breeding. Proceedings of the American Society for
Horticultural Science 71:412-415.
Gallagher, J.N. 1979. Field studies of cereal leaf growth. I. Initiation
and expansion in relation to temperature and ontogeny. Journal of
Experimental Botany 30:625-636.
Garcia-Huidobro, J., Monteith, J.L. and Squire, G.R. 1982. Time,
temperature and gennination of pearl millet (Pennisetum typhoides S.H.).
1. Constant temperature. Journal of Experimental Botany
33:288-96.
Gassnot, G. 1918. Boilrage zur physiologischer charaktcristik sommor und
winter annueller gewachse, insbesunder der Getreidepflanzen. Zeitschrift
fur Botanishe, 10:417-430).
Hiller, LK. and Kelly, W.C. 1979. Effects of post-vernalization temperature
on seedstalk elongation and flowering in carrots. Journal of American
Society of Horticultural Society, 104:257-262.
Hiller. L.K., Kelly, W.C. and Powell, I. E. 1979. Temperature interaction
with growth regulators and endogenous gibberellin-like activity during
seedstalk elongation in carrots. Plant Physiology 63:105 5-1061.
Monteith, J.L. 1981. Climatic variation and the growth of crops.
Quarterly Journal of the Royal Meteorological Society 197:749-74.
Ong, C.K. 1983. Response to temperature in a stand of pearl millet
(Pennisetum typhoides S. H.). I. Vegetable development. Journal
of Experimental Botany 34:322-36.
Robberts, E.H. and Summerfield, R.J. 1987. Measurement and prediction of
flowering in annual crops. In."Manipulation of Flowering". Atherton,
J.G. (Ed.), pp. 17-50. Butterworths, London.
Sakr, S. and Thompson, H.C. 1942. Effects of temperatures and photoperiod
on seedstalk development in carrot. Proceeding of the American Society
for Horticultural Science 41:343-346.
Copyright 1997 African Crop Science Society
|
| |||||||||

{kind=link}
{kind=link}