 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Some characteristics of pea necrosis elicitor from compatible interactions of Pseudomonas syringae p.v. pisi and Pisum sativum A.W. MWANG'OMBE^1 and S.A. ARCHER Department of Pure and Applied Biology, Imperial College of Science, Technology and Medicine. London. U. K. ^1 Department of Crop Science. University of Nairobi. P.O. Box 30197, Nairobi. Kenya. (Received 26 August 1993.: accepted 17 August 1995)
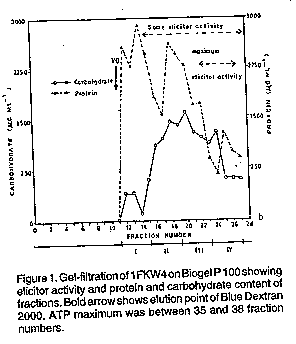
ABSTRACT Specific elicitors of pea necrosis were obtained from intercellular fluids of compatible race-cultivar interactions of Pseudomonas syringae pv. pisi and pea (Pisum sativum ). These elicitors induced necrosis in resistant but not in susceptible cultivars. Using pea cultivars carrying different resistance genes to the bacterial races 1 to 5 as test plants, an elicitor of necrosis was isolated and partially characterised. The necrosis inducing activity of these intercellular fluids was not lost after dialysis, gel-filtration and isoelectric focusing. The dialysed and lyophilised elicitor-active fraction of the intercellular fluids contained considerable amounts of protein and carbohydrate, and was partially sensitive to subtilisin BPN, proteinase K and protease V, but not to trypsin. These elicitors were inactivated by heat at 60 C and also by periodate. Most of the elicitor activity was recovered in a fraction of pH 11.7 after isoelectric focusing, suggesting a highly basic nature. Key Words: Carbohydrate, dialysis, gel filtration, isoelectric focusing, protein RESUME Des inducteurs specifiques de necrose ont ete obtenus des fluides intercellulaires d 'interactions compatibles de races de cultivars du Pseudomonas syringae pv.pisi et pois (Pisum sativum). Ces inducteurs declanchent la necrose dans les cultivars resistants mais pas dans ceux qui sont suceptibles. En utilisant comme plantes test des cultivars de pois ayant de genes de resistance aux races bateriennes 1 a 5, un inducteur de necrose a ete isole et partiellement caracterise. Les activites declanchees par la necrose dans les fluides intercellulaires n'ont pas ete perdues apres la dialyse, la filtration colloidale et la fixation isoelectrique. La dialyse et la fraction active lyophilisee et dialysee des inducteurs des fluides intercellulaires contenaient des quantites considerables de proteines et d'hydrates de carbone, et etaient partiellement sensible a la subtilisine BPN, a la proteinase K eta la protease V mais pas a la trypsine. Ces inducteurs etaient inactives par la chaleur a 60 C ainsi que par le periodate. La plupart des activivites des inducteurs ont ete recuperees dans une fraction de pH 11.7. Mots Cles: Carbohydrates, dialyse, infiltration de gel, ciblage isoelectrique, proteine INTRODUCTION Pseudomonas syringae pv pisi, the causal agent of pea bacterial blight, has several races (Taylor et at., 1984). In both the field and garden pea, there are several genes for resistance and gene-for-gene relationship has been proposed (Taylor et at., 1984). In liquid shake and still cultures growing on synthetic media, the bacterium produces elicitors which induce non-specific responses in resistant and susceptible pea cultivars (Mwang'ombe, 1987). However, intercellular fluids of compatible interactions of P. syringae pv. pisi and pea and culture filtrates, from membrane filter-sterilised medium containing susceptible pea stem homogenate inoculated with P. syringae pv. pisi, were shown to contain specific elicitors of necrosis (Mwang'ombe, 1987). The specificity of the elicitors was determined by the virulence genes present in the race of P. syringae pv. pisi (Mwang'ombe, 1987). Specific elicitors of necrosis have been detected in the intercellular fluids obtained l"rom compatible combinations of tomato and the tomato leaf mould pathogen, Cladosporium fulvum (de Wit and Spikman, 1982). Subsequent studies showed the elicitor of one race to be a low molecular weight (5500) peptide (de Wit et at., 1985). Beissman and co-workers (1992) isolated elicitor-active glycoproteins in apoplastic fluids of stem-rust infected wheat leaves. Pathogen-induced elicitors play cardinal roles in the initiation of recognition in incompatible host-pathogen interactions and some of the elicitors are host specific while others are non specific (Yoshikawa, 1983). This paper reports the isolation and partial characterisation of the elicitor of pea necrosis. A range of pea cultivars carrying different resistant genes to P. syringae pv. pisi were used as test plants for assaying necrosis induction. MATERIALS AND METHOD Plants, bacteria and inoculation. Pea cultivars carrying different genes for resistance to P. syringae pv. pisi were grown as described previously (Mwang'ombe, 1987). Cultures of P. syringae pv. pisi were grown on King's B medium (King et al., 1954). Stems of 12-14 days old cv. Kelvedon Wonder seedlings were inoculated with a bacterial suspension of 10^9 colony forming units (CFU) using a modified method of stem inoculation (Mwang'ombe, 1987). Isolation of elicitors and assay of the biological activity. Stems of the susceptible cultivar (cv. Kelvedon Wonder) which showed visible macroscopic symptoms (9 days after inoculation ), were routinely harvested for the isolation of elicitors. Stems were infiltrated with sterile distilled water in vacuo and the intercellular fluids isolated and sterilised (Mwang'ombe, 1987). The fluids were used immediately or stored at -20 C. Pea seedlings, 12-14 days old, were injected with intercellular fluids using a syringe and then incubated. Cultivars (cvs) Kelvedon Wonder, Fortune, and Feltham First were routinely used as test plants for assaying the induction of necrosis during purification and characterisation of the elicitor. However, in some experiments, other cultivars were also used viz. Early Onward, Meteor, Sprite, Puget, Hurst Green Shaft and Partridge. The pea cultivars carried different genes for resistance to P. syringae pv. pisi. Purification of elicitors. Gel filtration was used for initial purification of the necrosis-inducing elicitor. Bio-Gel P 100(100-200 mesh)from BioRad was prepared according to the manufacturer's instructions. Bio-Gel P 100(15 g) was gradually added to sterile potassium phosphate buffer (pH 7.5, 0.05M) containing 0.1 M KCl, hydrated for four hrs at room temperature then degassed using a suction pump. The gel suspension was allowed 10 reach 4-5 C before packing was started. The column was connected 10 the buffer reservoir and eluted using a peristaltic pump (Minipuls 2-Gilson, Anachem). Flow rate was 12 ml hr and fractions of 4 ml were collected. Blue Dextran 2000 and ATP were used to establish the void volume and total volume of the column, respectively. The former was detected by monitoring of the collected fractions at 650 nm and the latter at 260 nm. This column was used to resolve 5 ml samples of filter sterilised intercellular fluid, from cv. Kelvedon Wonder inoculated with race 4, previously lyophiliscd to half volume. Following the void volume, determined to be 44 ml using Blue Dextran 2000, 4 ml fractions were collected which were grouped into 12 sets of four fractions. Fractions from gel-filtration were dialysed in visking tubing (Medice/Int. Ltd.) against repeated changes of distilled water at 4-5 C. Dialysis tubing was pre-treated according to Plummer (1978) The dialysed fractions were freeze-dried and then dissolved in 2.5 ml sterile distilled water. The fractions were assayed using leaves of the cultivars named above. Protein estimation. Protein was determined using the Folin-Ciocalteu phenol reagent (Lowry et al.,1951) with bovine serum albumin (BSA) of up to 500 ug ml^-1 as the standard. To a BSA or intercellular fluid sample of 0.5 ml, 3 ml of reagent C (50 ml of reagent A plus 1 ml of reagent B; reagent A being 2% Na2CO3 in 0.1M NaOH and reagent B was 0.5% CuSO4. 5H2O in 1% potassium (+) tartarate) was added, mixed well and allowed to stand for 20 minutes at room temperature. Finally, 0.3 ml of reagent E (Folin Ciocalteu phenol reagent (BDH) solution diluted twofold to approximately 1 N in acid) was added rapidly and mixed within a second or two. After 30 minutes or longer, the absorbance of the samples was determined using a spectrophotometer (Beckman series 30 UV-visible) at 750 nm. Blanks contained 0.5 ml of 0.1 M NaOH instead of protein. Determination of carbohydrates. The phenolsulphuric acid method (Dubois et al., 1956) with slight modification, was used for carbohydrate determination and estimation using glucose at up to 500 ug ml^-1 as standard. To 0.5 ml of glucose or intercellular fluid sample solution, 0.5 ml of 5% (w/v) phenol in sterile distilled water was added. Finally, 2.5 ml of concentrated sulphuric acid was added rapidly; the stream of acid directed against the liquid surface rather than against the side of the test tube to give good mixing. The test tubes were allowed to stand for 10 min at room temperature, then shaken and placed for 20 min in a water-bath at 30 C before the absorbence at 490 nm was measured against a water blank. Effect of heat on the hypersensitive reaction - inducing activity of intercellular fluid. Stability to heat was tested by incubating crude intercellular fluid (before Bio-Gel P 100 filtration) for 15 minutes at the following water-bath temperatures, 30, 40, 50, 60, 70, 80, 90 and 100 C. The samples were assayed for induction of symptoms using cvs Fortune, Feltham First and Kelvedon Wonder. Untreated intercellular fluid was included as a control. Effect of proteases on intercellular fluid hypersensitive reaction-inducing activity. Sensitivity to proteases was tested by incubating intercellular fluid pooled fractions after gel filtration in an aqueous solution containing 1 mg ml^-1 subtilisin BPN, proteinase k, protease V or trypsin for 24 hr at 37 C. The mixtures were assayed on leaves of cvs Fortune, Kelvedon Wonder and Feltham First, together with controls of enzyme or intercellular fluid alone. An earlier experiment (not reported here) showed that the temperatures at which proteases were inactivated were higher than that which inactivated the elicitor. Thus, heat inactivation could not be used as a method of removing the effects of proteases from the mixtures before assay. This experiment was repeated using subilisin BPN only at 3 mg ml^-1. Periodate oxidation of intercellular fluids. Sensitivity to HIO4 was tested by incubating the intercellular fluid in periodic acid in sodium acetate buffer. One ml of 0.05 M periodic acid in 0.1 M sodium acetate buffer, pH 4.8, was thoroughly mixed with 1 ml of sample and incubated at 20 C in the dark for 8 hr. The sample mixture was then dialysed extensively against distilled water, with four changes at 4-5 C, to remove excess periodate and sodium acetate. The sample was assayed on cvs Fortune, Feltham First and Kelvedon Wonder. Isoelectric fractionation and characterization of intercellular fluid. Isoelectric focusing in natural pH gradients was used to separate the proteins in intercellular fluid by their isoelectric points using the method of Vesterberg and Svensson (1966). An electrofocusing column of 110 ml capacity with cooling mantle and electrodes (LKB 8101) was used. Cold water at 5 C from a thermostated bath was circulated through the mantle. The electric power supply (LKB type number 3371E) which could give up to 1200 V and 50 mA was used. The ampholytes used were pharmalyte pH 3-10 from Pharmacia. Phosphoric acid was used as the anode electrolyte which was arranged at the base of the sucrose density gradient. Sodium hydroxide comprised the cathode solution and carrier ampholytes were used at 2% (V/V). After focusing for 48 hr, 4 ml fractions were collected, measured for pH, exhaustively dialysed against distilled water, lyophilised to 2 ml and bioassayed for intercellular fluid activity. Protein and carbohydrate were estimated on 10^2 dilutions of each fraction. RESULTS Purification of intercellular fluid by gel filtration. Tests using sets bulked in pairs showed that sets I to IV (Fig. 1) induced extensive light brown lesions when infiltrated into leaflets and stems of cvs Fortune, Feltham First, Partridge and Hurst Green Shaft, all of which are resistant to race 4. The characteristic narrow, dark green, water-soaked margin of the necrotic lesions, typical of intermediate resistance, was observed in the latter two cultivars. More extensive water soaking associated with the susceptible reaction was induced in cvs Kelvedon Wonder, Early Onward and Puget. Fractions in sets V and VI induced a general chlorosis in all cultivars tested. Other fractions in sets VII-XII did not induce symptoms in all cultivars tested. Further bioassay using seven cultivars, Fortune, Feltham First, Partridge, Hurst Green Shaft, Kelvedon Wonder, Early Onward and Puget revealed that elicitor activity resided in sets II to IV with a peak in set IV (Fig. 1 ). Attempts to localise elicitor activity further by assaying every 4 ml fraction on cvs Fortune, Feltham First and Kelvedon Wonder, were not successful despite the activity in the pooled sets being detectable down to a dilution of 1:8 (data not reported here). The peak of elicitor activity did not correspond with the peak of either protein or carbohydrate, although there was an appreciable component of both in all active fractions (Fig. 1). The total protein and carbohydrate contents of sets 1 to IV inclusive were 117 mg and 60 mg, respectively, representing approximately 15% of crude intercellular fluid from cv. Kelvedon Wonder inoculated with race 4.
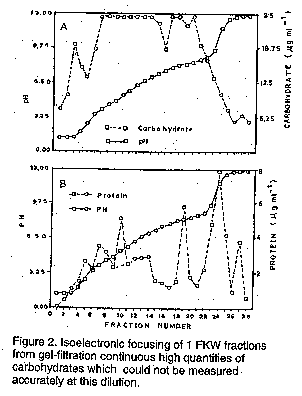
Sensitivity of intercellular fluids to periodate and heat. Sets II to IV from gel filtration of intercellular fluid lost their elicitor activity after treatment with sodium periodate for 8 hr. Infiltrated leaflets of cvs Fortune, Feltham First and Kelvedon Wonder remained symptomless. The control treatment of sodium periodate in acetate buffer was extremely phytotoxic to all pea cultivars tested. Heat treatment of intercellular fluid for 15 minutes at temperatures up to 50 C had no effect on the intercellular biological activity when assayed on cvs Fortune, Feltham First and Kelvedon Wonder. At 60 C most activity was lost although minute light brown lesions were still formed on cv Feltham First. Generally higher temperature destroyed activity. Isoelectric focusing of intercellular fluid fractions from gel filtration. Bulked material from sets II to IV from several gel filtration experiments was subjected to isoelectric focusing in a sucrose density gradient. Fraction 24 possessed the greatest biological activity, inducing extensive necrosis in cvs Fortune, Feltham First, Partridge and Hurst Green Shaft within 48 hr. In contrast, the race 4 - susceptible cultivars Kelvedon Wonder, Early Onward and Puget were symptomless 48 hr after infiltration, although by the seventh day very small discoloured lesions had appeared at the point of injection. The adjacent fraction, 23, was the only other to possess any (weaker) necrosis inducing activity. Isoelectric focusing effectively separated the necrosis-inducing factor from other substance(s) causing water soaking. This latter ability was found in fractions 7 to 17 each of which induced water-soaked symptoms, mainly on the abaxial surface, without tissue death. The spectrum of protein and carbohydrate (Fig.2a, b) reveals that fraction 24, which had the greatest biological activity, also contained most protein. The pH 11.7 of this fraction is deceptive since this coincided with a very steep region of the pH gradient as evidenced by the adjacent fraction 23 having a lower pH. Fraction 24 had a relatively low carbohydrate content, most of which was present in fractional values off scale in the phenol- sulphuric acid assay even at 1:100 dilution. DISCUSSION
The necrosis-inducing activity of the elicitors in this study was not lost after dialysis, gel filtration and isoelectric focusing. The elicitor was sensitive to HIO4 and was heat inactivated at 60 C. The elicitor was partially sensitive to subtilisin proteinase K and protease V but not to trypsin at the concentration tested. Most of the elicitor was recovered at a high pH fraction after isoelectric focusing suggesting highly basic properties. Exogenous inhibitors of hypersensitive reaction have been reported before (Sasser, 1978; Keen et al., 1981; de Wit et al., 1985; Beissman et al., 1992). The elicitor isolated by de Wit and co-workers (1985) was a small peptide sensitive to pronase and protease. Beissmann et al. (1992) reported that the elicitor- activity of the intercellular washing fluid glycoprotein resides in the carbohydrate moiety because periodate, not trypsin or pronase, destroyed activity. Although the results of this study are not conclusive, we suspect that the elicitor(s) of necrosis in intercellular fluids of compatible race-cultivar interactions could be a glycoprotein. Keen et al. (1981) reported that blasticidin S inhibited hypersensitivity reaction in an incompatible soybean - P. syringae pv. glycinea combination allowing the multiplication of the bacteria and development of typical water-soaked disease symptoms.
The cloning of avirulence genes of P. syringae pv. glycinea (Staskawicz etal., 1984; Staskawicz et al., 1985) and Xanthomonas campestris pv. malvacearum (Gabriel, 1985) as well as the occurrence of a similar gene(s) in P. syringae pv. pisi (Taylor et al., 1984) and on a conjugative plasmid in X. campestris pv. vesicatoria (Stall et al., 1984) constitute highly significant developments in understanding the molecular basis for disease resistance and host specificity. De Wit etal. (1986) suggested that the race-specific elicitor of necrosis from C. fulvum is the direct product of an avirulence gene. This may also be true for the P. syringae pv. pisi elicitors in this study. If this is the case then these studies constitute an important link between the molecular genetics and physiology of resistance. Therefore, the isolation of such molecules (elicitors) could be used to provide more information on cellular, physiological and biochemical interactions of hosts and pathogens, and thus help further to unravel the mystery of recognition mechanism in host specificity. ACKNOWLEDGEMENTS This paper is a portion of a Ph.D. thesis submitted by the first author to the Imperial College of Science, London, UK. REFERENCES Beissmann, B. Engels, W., Kogel, K., Marticke, K.H. and Reisener, H.J. 1992. Elicitor-active glycoproteins in apoplastic fluids of stemrust-infected wheatleaves. Physiologicaland Molecular Plant Pathology 40:79-89. De Wit, P.J.G.M. and Spikman, G. 1982. Evidence for the occurrence of race- and cultivar-specific elicitors of necrosis in intercellular fluids of compatible interactions of Cladosporium fulvum and tomato. Physiological Plant Pathology 21:1 - 11. De Wit, P.J.G.M, Hofman, A.E., Velthuis, G.C.M. and Kuc, J.A.1985. Isolation and characterization of an elicitor of necrosis isolated from intercellular fluids of compatible interactions of Cladosporium fulvum (syn. Fulvia fulva) and tomato. Plant Physiology 77:642-647. De Wit, P.J.GM, Buurlage, M.B. and Hammond, K.E. 1986. The occurrence of host-pathogen and interaction-specific proteins in the apoplast of Cladosporium fulvum (Syn. Fulviafulva) infected tomato leaves. Physiological and Molecular Plant Pathology 29:159-172. Dubois, M., Gilles, K.A., Hamilton, J.K. Rebers, P.A. and Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Analytical Chemistry 28:350356. Ellingboe, A.H. 1984. Genetics of host.-parasite relations: An essay. In: Advances in Plant Pathology. Ingram, D.S. and Williams, P.H. (Eds.), 2:131 - 15 1. Academic Press, London. Gabriel, D .W. 1985. Molecular cloning of specific avirulence genes from Xanthomonas campestrispv. malvacearum. In: Advances in the Molecular Genetics of the Bacteria Plant Interaction. Szalay, A.A. and Legocki, R.P. (Eds.), pp. 202-204. Media Services, Cornell University, Ithaca, New York. Keen, N.T., Ersek, T., Long, M., Bruegger, B. and Holliday, M. 1981. Inhibition of the hypersensitive reaction of soybean leaves to incompatible Pseudornonas spp. by blasticidin S, streptomycin or elevated temperature. Physiological Plant Pathology 18:325-337. King, E.O., Ward, M.K. and Raney, D.E. 1954. Two simple media for the demonstration of pyocyanin fluorescein. Journal of Laboratory and Clinical Medicine 44:301-307. Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J. 1951. Protein measurement with the Folin phenol reagent. Journal ofBiological Chemistry CXCIII. 265-275. Mwang'ombe, A.W. 1987. Specificity in Pea Blight Disease Caused by Pseudomonas syringae pv. pisi. Ph.D. Thesis, University of London. 159pp. Plummer, D.T. 1978. An Introduction to Practical Biochemistry. 2nd edition, pp. 47-53. McGraw-Hill, London. Sasser, M. 1978. Involvement of bacterial protein in induction of the hypersensitive reaction in tobacco. Phytopathology 68:361 -363. Stall, R.E., Loschke, D.C. and Rice, R.W. 1984. Conjugational transfer of copper resistance and avirulence to pepper within strains of Xanthomonas campestris pv. vesicatoria. Phytopathology 74:797 (Abstr). Staskawicz, B.J., Dahlbeck, D. and Keen, N.T. 1984. Clones avirulence gene of Pseudomonas syringae pv. glycinea determines race-specific incompatibility on Glycine max.(L.) Proceedings of the National Academy of Sciences (USA) 81:6024-6028 Staskawicz, B.J., Dahlbeck, D., Napoli, C. and Keen, N.T. 1985. Molecular genetics of plantbacterial pathogen specificity. In: Advances in the Molecular Genetics of the Bacteria Plant Interaction. Szalay, A.A. and Legocki, R.P. (Eds.), pp. 195-198. Media Services, Cornell University Press, Ithaca, New York. Taylor, J.D., Vivian, A. and Malik, A. 1984. The genetics of Pseudomonas syringae pv. pisi. In: Proceedings of 2nd Working Group of Pseudomonas syringae pathovars. Panagopoulos, C.G., Psallidas, P.G. and Alivizatos, A.S. (Eds.), pp. 90-91. Hellenic Phytopathological Society, Athens. Vesterber, O. and Svensson, H. 1966. Isoelectric fractionation, analysis and characterization of ampholytes in natural pH gradients. Acta Chemica Scandinavica 20:820-834. Yoshikawa, M. 1983. Macromolecules, recognition and triggering of resistance. In: Biochemical Plant Pathology. Callow, J.A. (Ed.), pp. 267-298. John Wiley and Sons, New York. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Line drawing images[cs96044b.gif] [cs96044a.gif] |
| |||||||||

{kind=link}
{kind=link}