 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Identification and characterisation of a potyvirus of Bambara groundnut
DIJKSTRA, E. KEESEN, M. BRINK^1, D. PETERS and H. LOHUIS
Department of Virology, Wageningen Agricultural University,
Binnenhaven 11,6709 PD Wageningen, The Netherlands (Received 12 April, 1995; accepted 6 November, 1995)
Code Number: CS96045
Sizes of Files:
Text: 26K
Graphics: Photographs (jpg) - 109KABSTRACT A potyvirus was isolated from Bambara groundnut (Vigna subterranea (L.) Verdc.) plants raised in a glasshouse from seeds obtained from Zimbabwe. Infected plants showed stunting and foliar discolouration. The cause of the disease was identified to be a strain of peanut mottle virus. This was based on host reaction, serology, properties in crude sap, type of cylindrical inclusions and transmission characteristics. The strain was, consequently, named the Bambara groundnut strain of peanut mottle virus. Key Words: Bean black root virus, bean common mosaic virus, cowpea aphid-borne virus. peanut mottle virus RESUME Un potyvirus etait isole des plants d 'arachide de Bambara (Vigna subterranea) eleves sous serre vitree. Ces plants provenaient de semences obtenues de Zimbabwe. Les plants infectes montraient un arret de croissance et une decoloration foliaire. La cause de la maladie etait identifiee comme etant la fatigue due au virus de marbrure d'arachide. Ceci etait du a la reaction de l'hote, a la serologie, aux proprietes de la seve brute, au type d'inclusions du cylindre et de caracteristiques de transmission de la maladie. La race etait par consequent denommee virus de marbrure d'arachide de Bambara. Mots Cles: Virus de racine noire d'haricot, virus de la mosaique commune, virus du niebe transmis par les aphides, virus de marbrure d'arachide INTRODUCTION Bambara groundnut (Vigna subterranea), a leguminous plant, is grown as an indigenous crop in arid tropical Africa for seed and forage. Its generic name was Voandzeia, but was renamed Vigna in 1980 (Limeman and Azam Ali, 1993). During phenological studies at the Department of Agronomy of Wageningen Agricultural University on plants of Bambara groundnut raised from seeds harvested in Zimbabwe, seedlings were occasionally observed to have an abnormal appearance consisting of stunting (Figure 1), dark green mosaic and sometimes chlorotic mottling. "Chop" preparations for the electron microscope of these plants showed the presence of potyvirus like particles. Four potyviruses are known to occur in Bambara groundnut, viz. Bambara groundnut mosaic virus (Bird and Corbett, 1988), cowpea aphid-borne mosaic virus (Ng et al., 1985), peanut mottle virus (Bock, 1973; Bock et al., 1978; Li et al., 1991) and Voandzeia distortion mosaic virus (Fauquet and Thouvenel, 1987; Brunt et al., 1990 ). The present paper deals with the identification and further characterisation of the causal agent of Bambara groundnut disease occurring in Zimbabwe. MATERIALS AND METHODS
Virus isolates and strains. The virus from Bambara groundnut was isolated by mechanical transmission from diseased plants to Nicotiana benthamiana in which it was maintained and propagated. For comparison, the following potyviruses were used: bean black root virus (BBRV) strain NL3, formerly considered to be a strain of bean common mosaic virus (BCMV), BCMV strains blackeye cowpea - Indonesia (BIC/Ind), blackeye cowpea - Wageningen (BIC/W) (formerly called blackeye cowpea mosaic virus (BICMV) strains Ind and W, respectively) and NY15, and cowpea aphid-borne mosaic virus strain-Morocco (CAMV-Mot), formerly considered to be a strain of BICMV (Dijkstra et al., 1987; Lana et al., 1988; Khan et al., 1990). Test plant reactions. In host range tests, three plants of each of 35 species and cultivars were inoculated with diluted crude sap from virus-infected N. benthamiana plants using Carborundum (600 mesh) as the abrasive. After three to four weeks, presence or absence of infection was checked by back inoculation to Chenopodium amaranticolor. Properties in crude sap. Sap from infected leaves of N. benthamiana was used as inoculum and assayed on plants of C. amaranticolor in experiments to determine the dilution end-point, thermal inactivation point and longevity in vitro of the virus. Transmission by aphids. Transmission experiments were carried out with apterous aphids (Myzuspersicae) from virus-free cultures. Aphids were starved for 60 rain and then given an acquisition access period on young diseased plants of N. benthamiana for about 10 min. The aphids were then transferred to young N. bentharniana plants for an inoculation feeding period of about 20 min (10 or 14 plants with 10 aphids per plant). To prevent possible mechanical transmission of virus with the paint-brush used for aphid transfer, small pieces of paper were placed on the leaves of assay plants and the aphids were transferred on to them. Eventually, the aphids were killed with an insecticide. Presence of virus in the assay plants was established by symptomatology and dot immunobinding assay.
The ultraviolet-light absorption of the virus suspension was determined using an extinction coefficient of 2.4, as for tobacco etch virus (Purcifull, 1966). For determination of buoyant density, equilibrium centrifugation of purified virus was carried out in CsCl solution, prepared by dissolving 2 g of CsCl in a mixture of 100 ml (or 400 ml) virus suspension and Tris/HCl buffer to a final volume of 5.5 ml, at 38,000 rpm for 17 hr. Thereafter, 0.5 ml fractions were collected, the position of the virus established and the refractive indices of the fractions determined with a refractometer. The buoyant density of the virus was calculated from the refractive indices of collected fractions, using tables relating refractive index and density at 25 C (Anderson and Anderson, 1973). Electrophoresis of the capsid protein. The relative molecular mass (Mr) of the capsid protein was determined by sodium dodecyl sulphate (SDS)-polyacrylamide gel electrophoresis on a vertical 12% gel with a 4% stacking gel (Laemmli, 1970) in a BioRad Mini Protean II system. A purified virus suspension mixed 1:1 with 0.06 M Tris/HCl, pH 6.8, containing 10% (v/v) glycerol, 2% (w/v) SDS, 2-b-mercapto-ethanol, and 0.00125% (w/v) bromophenol blue, was boiled for 3 min before electrophoresis at 150 V constant. Gels were fixed in 10% (v/v) methanol, and destained with 7% (v/v) acetic acid and 5% (v/v) methanol. Phosphorylase B (M[r] 94,000), bovine serum albumin (M[r] 67,000), ovalbumin (M[r] 43,000), carbonic anhydrase (M[r], 30,000), soybean trypsin inhibitor (M[r] 21,000) and o-lactalbumin (M[r] 14,400) were used as M[r] markers. Serology. For the production of an antiserum, a mixture of 1 ml of a purified virus suspension (0.2 mg ml^-1) and 1 ml of Freund's incomplete adjuvant was injected intramuscularly into a rabbit. After 14 days, a second injection was given likewise with 0.5 mg ml^-1 virus and emulsified with Freund's incomplete adjuvant. Three weeks later, blood was collected for the first time and then after two weeks. The titre was determined by microprecipitation under paraffin oil. Dilution series of antiserum and antigen were prepared with O. 1 M Tris-citric acid at pH 8.0. The antiserum to peanut mottle virus (PeMoV) was provided by Mr. S.E. Albrechtsan (Copenhagen, Denmark) who obtained it from Dr. D.V.R. Reddy (ICRISAT, Hyderabad, India). To prevent non-specific reactions, an absorption step with sap from healthy N. benthamiana or Phaseolus vulgaris plants (diluted 1:20 in PBS-Tween) was carried out for all antisera, except the one to virus from Bambara groundnut from which no background was obtained when testing healthy plant extracts. Serological relationships with other potyviruses were studied in indirect ELISA using the direct antigen coating procedure adopted from Hobbs et al. (1987). Absorbance was recorded after 10 and 30 min with an ELISA microplate reader at 405 nm. Dot immunobinding assay was carried out as described by Hibi and Saito (1985) for dot-blot immunoassay. Each sample tested consisted of clarified sap from leaves of N. benthamiana mixed 1:9 with PBS Tween. Light microscopy. Epidermal strips of primary leaves of Vigna unguiculata were prepared for light microscopy using Azure A and "Luxol" brilliant-green BL-calcomine orange 2RS (O-G) stains (Christie and Edwardson, 1977). The strips were mounted in Euparal and examined in a Leitz Orthoplan microscope. Electron microscopy. Carbon-reinforced formvar-coated copper grids (400 mesh) were placed downwards on the surface of droplets of purified virus for two min, stained with 2% (w/v) uranyl acetate and examined with a Philips CM 12 transmission electron microscope. For calculating virus particle length, a comparison was made with micrographs of catalase-coated grids, using the periodicity of catalase crystals (8.6 + 0.2 nm). To this end, a droplet of catalase crystal suspension was placed on a grid and after a few rain, replaced by a drop of 2% (w/v) phosphotungstate at pH 5.5, for 15 sec. Thereafter, the grid was dried and examined. "Chop" preparations were prepared as follows. A diseased leaf was chopped in a drop of distilled water till the latter turned light green. The drop was placed on a carbon-reinforced formvar-coated copper grid for about one min and then slowly removed by blotting. Staining was done with 2% (w/v) phosphotungstate at pH 5.5 for 15 sec. TABLE 1. Reactions of test plants to a potyvirus isolate from Bambara groundnut (Vigna subterranea)
Reactions
_____________________
local systemic
_________________________________________________________
Arachis hypogaea + ++
Beta vulgaris 'Groeningia' ++ ++
Capsicum annuum 'Westlandse Zoete' ++ ++
Chenopodium amaranticolor CS, NS ++
Chenopodium quinoa CS ++
Crotalaria juncea + +
Cucumis sativus 'Lange Groene Gigante' ++ ++
Datura stramonium ++ ++
Gomphrena globosa ++ ++
Lycopersicon esculentum 'Moneymaker' ++ ++
Nicotiana benthamiana # LC, LD, Mot, VC
Nicotiana clevelandii # Mos, Mot
Nicotiana glutinosa ++ ++
Nicotiana occidentalis CS, NS C, NS, ##
Nicotiana rustica + +
Nicotiana tabacum 'Samsun' + ++
Nicotiana tabacum 'White Budey' + ++
Ocimum basilicum ++ ++
Petunia hybrida + ++
Phaseolus vulgaris 'Bataaf' CS ++
Phaseolus vulgaris 'Dubbele Witte' NS Mot
Phaseolus vulgaris 'Pinto' NS ++
Physalis floridana ++ ++
Pisum sativum 'Kelvedon Wonder' + +
Solanum melongena 'Lange Violerie' ++ ++
Spinacia oleracea ++ ++
Tetragonia expansa ++ ++
Trifolium repens ++ ++
Vicia faba + ++
Vigna cylindrica # Mos
Vigna radiata ++ ++
Vigna subterranea + Mos, Mot, St
Vigna unguiculata 'California Black,eye'
CS, NS, VN Mos, VB
Vigna unguiculata 'Early Red' ++ ++
Zinnia elegans ++ ++
______________________________________________________________C = chlorosis; CS = chlorotic spots; LC = ear curling; LD = leaf deformation; Mos = mosaic; Mot = mottling; NS = necrotic spots; St = stunting; VB = vein banding; VC = veinal chlorosis; VN = veinal necrosis; # = no symptoms; ++ = no infection; + = symptomless infection; ## = plant dead. RESULTS Host range. The virus was readily sap transmissible from diseased Bambara groundnut plants to N. benthamiana and Chenopodium spp. causing veinal chlorosis, mottling and leaf curling in the former and local lesions in the latter. Of the 35 species and cultivars tested, 11 showed symptoms and eight were symptomlessly infected (Table 1). Purified virus suspensions inoculated to young and healthy Bambara groundnut seedlings, caused symptoms similar to those of the original diseased plants. Properties in crude sap. The virus in crude sap from infected N. benthamiana plants had a dilution end-point of 10^2 - 10^-'3, a thermal inactivation point of 50 - 60 C, and infectivity retained at 24 C for 10 - 12 days. Transmission by aphids. Myzus persicae transmitted the virus to 4 of 14 N. bentharniana plants, to 2 of 10 plants and to 2 of 10 plants in the three experiments, respectively. Virus purification. Virus yields ranged from 1.5 to 2.0 mg per 100 g of infected N. benthatniana leaves.
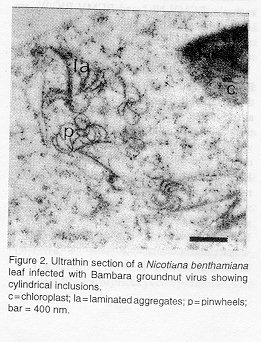
Properties of particles. Purified virus suspensions had absorption maxima and minima at 262 and 245 nm, respectively, and the A260/A280 ratio was between 1.21 and 1.29 without correction for light-scattering. The buoyant density in CsCl was 1.30-1.31 g cm^-3. Polyacrylamide gel electrophoresis of the coat protein revealed one band with an estimated M[r] of c. 37,000 Da. Serology. The antiserum to the virus from Bambara groundnut had a homologous titre of 1:128 in microprecipitation tests. In ELISA, both diluted virus-containing sap and purified virus preparations cross-reacted strongly with antiserum to PeMoV. A less strong reaction was observed with antiserum to NYIS, but the latter was nonreciprocal (Table 2). None of the antisera to other members of the BCMV-subgroup (BIC/Ind, BIC/ W, NL3, Mor) gave positive reactions with tile virus from Bambara groundnut. Light microscopy. O-G stained specimens showed masses of cylindrical inclusions in a number of epidermal cells of primary leaves of V. unguiculata with diffuse chlorotic rings. Electron microscopy. The flexuous virus particles had a normal length of 806 nm +/- 0.025%. Ultrathin sections of infected leaves of N. benthamiana showed pinwheels, scrolls and laminated aggregates (Fig. 2), typical of representatives of subdivision III of potyviruses (Edwardson, 1974).
A seed-transmissible potyvirus causing mosaic symptoms in Bambara groundnut has been reported from West Africa (Bird and Corbett, 1988). Bock (1973) and Bock et al. (1976, 1977. 1978) had previously described a potyvirus isolated from V. subterranea in East Africa which was serologically related to PeMoV, but differed from it in being able to infect C. amaranticolor. It was. therefore, considered to be a distinctive host-adapted strain of PeMoV; this isolate, however. did not infect groundnut (Book et al., 1977, 1978). The virus isolate from Bambara groundnut described here, is serologically closely related to PeMoV. It causes local lesions in Chenopodium spp., thus resembling the above-mentioned strain of PeMoV and only differing from the latter in causing symptomless local infection in groundnut (Arachis hypogaea). Besides the above-mentioned reactions on Chenopodium spp. and the symptomless infection of groundnut, the virus further differed in host range from the type strain of PeMoV (Brunt et al., 1990) especially in its infectionability of Crotalaria juncea (locally and systemically), N. rustica (locally and systemically), N. tabacum and petunia (locally) and its inability to infect Cucumis sativus. It also differed from the VS strain of PeMoV (Li et al., 1991 ) in reactions on Chenopodium spp. as well as the symptomless infections of groundnut, Nicotiana spp., Pisum sativum and Vicia faba. With respect to its properties in crude sap, seed and aphid transmission, and type of inclusion bodies (pinwheels, scrolls and laminated aggregates) Bambara groundnut virus resembles the type strain of PeMoV (Bock and Kuhn, 1975) to which it is serologically closely related, as stated earlier. It is distantly related to BCMV-NY15, but has no serological relationship with BCMV-BIC (former BICMV), as reported for PeMoV (Rajeshwari etal. 1983) and also not with CAMV-Mot. The estimated M of the capsid protein is higher (37,000 Da) than the one reported for PeMoV (34,000 Da; and for PeMoV-VS (31,000-32,000 Da) (Li et al., 1991). However, sometimes even strains of the same virus, for example, BCMV-NL1 and BCMV-BIC/W, are known to possess different values of M for their capsid proteins, viz. 39,000 and 36,000 Da, respectively (Lana et al., 1988). The virus isolated from Bambara groundnut by Bird and Corbett (1988) appears to be identical to the virus reported here, but no proper comparisons are possible due to the incomplete description of the virus. Identity of Voandzeia distortion mosaic virus can be ruled out, because of the latter's lack of seed transmissibility, different symptoms on Bambara groundnut and no serological relationship with PeMoV. Based on the results obtained in the present study, it is concluded that the virus isolated from Bambara groundnut plants is a strain of PeMoV which could be designated the Bambara groundnut strain of peanut mottle virus. TABLE 2. Comparison of absorbancies (A[405]) in ELISA of the Bambara groundnut (Ba) virus with bean common mosaic virus strains blackeye cowpea - Indonesia (BIC/Ind), blackeye cowpea - Wageningen (BIC/W) and NY15, bean black root virus strain NL3 (NL3), cowpea aphid-borne mosaic virus strain Morocco (Mor) and peanut mottle virus (PeMoV)
Antisera Ten times diluted sap of healthy (H) Purified
plants and those infected with tested
suspens-
ion^1
--------------------------------------------------------------
H Ba BIC/Ind BIC/W NY15 NL3 Mor Ba
--------------------------------------------------------------
Ba 0.045 0.633 0.020 0.000 0.011 0.033 0.021 0.765 BIC/Ind
0.040 0.002 1.160 0.599 0.881 0.160 0.138 0.043 BIC/W 0.046
0.006 0.371 0.370 0.484 0.330 0.106 0.049
NY15 0.049 0.260 1.358 1.220 1.559 1.144 0.776 0.295
NL3 0.034 0.000 0.232 0.180 0.278 0.424 0.192 0.040
Mor 0.125 0.076 0.235 0.233 0.000 0.195 0.586 0.096
PeMoV 0.134 0.472 0.400 0.254 0.297 0.213 0.341 0.493^1 Concentration of 400 ng ml^-1 ACKNOWLEDGEMENTS We thank Mr S.E. Albrechtsen (Copenhagen, Denmark) for sending antiserum to peanut mottle virus and for sharing his experience with us. REFERENCES Anderson, N.G. and Anderson, N.L. 1973. Selected data molecular biology. CRC Press, Cleveland, U.S.A. Bird, A.E. and Corbett, M.K. 1988. Bambarra groundnut mosaic caused by a seed-transmitted potyvirus. Phytopathology 78:860. Bock, K.R. 1973. Peanut mottle in East Africa. Annals of Applied Biology 74:171 - 179. Bock, K.R. Guthrie, E.J. and Meredith, G. 1977. 750 nm virus isolates from Voandzeia, Cassia and Phaseolus lunatus. In: Record of Research for the period 1st January to 31st December 1975. Annual Report 1975. East African Agriculture and Forestry Research Organisation, Nairobi, Kenya: 122-123. Bock, K.R. Guthrie, E.J. and Meredith, G. 1978. Viruses occurring in East Africa that are related to peanut mottle virus. Annals of Applied Biology 89:423-428. Bock, K.R., Guthrie, E.J, Meredith, G. and Njuguna, J.G.M. 1976. Viruses of other leguminous crop plants. In: Record of Research for the period 1st January to 31st December 1974. Annual Report 1974. East African Agriculture and Forestry Research Organization, Nairobi, Kenya: 125-126. Bock, K.R. and Kuhn, C.W. 1975. Peanut mottle virus. CMI/AAB Descriptions of Plant Viruses. No. 141:4 pp. Brunt, A. Crabtree, K. and Gibbs, A. 1990. Viruses of Tropical Plants. C.A.B. International, Wallingford, U.K. Christie, R.G., and Edwardson, J.R. 1977. Light and electron microscopy of plant virus inclusions. Florida Agricultural Experiment Stations Monograph Series 9, Gainesville, U.S.A.: 155 pp. Dijkstra, J., Bos, L., Bouwmeester, H.J., Hadiastono, T. and Lohuis, H. 1987. Identification of blackeye cowpea mosaic virus from germplasm of yard-long bean and from soybean, and the relationships between blackeye cowpea mosaic virus and cowpea aphid-borne mosaic virus. Netherlands Journal of Plant Pathology 93:115-133. Edwardson, J.R. 1974. Some properties of the potato virus Y-group. Florida Agricultural Experiment Stations Monograph Series 4, Gainesville, U.S.A.: 398 pp. Fauquet, C. and Thouvenel, J .C. 1987. Voandzeia distortion mosaic potyvirus. In: Plant Viral Diseases in the Ivory Coast. ORSTOM Paris, Documentation Techniques No. 46:243 pp. Hibi, T. and Saito, Y. 1985. A dot-blot immunobinding assay for the detection of tobacco mosaic virus in infected tissue. Journal of General Virology 66:1191-1194. Hobbs, H.A., Reddy, D.V.R., Rajeshwari, R. and Reddy, A.S. 1987. Use of direct antigen coating and protein A coating ELISA procedures for detection of three peanut viruses. Plant Disease 71:747-749. Khan, J.A., Lohuis, H. Goldbach, R.W. and Dijkstra, J. 1990. Distinction of strains of bean common mosaic virus and blackeye cowpea mosaic virus using antibodies to N- and C- or N-terminal peptide domains of coat protein. Annals of Applied Biology 117:583-593. Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685. Lana, A.F., Lohuis, H., Bos, L. and Dijkstra, J. 1988. Relationships among strains of bean common mosaic virus and blackeye cowpea mosaic virus - members of the potyvirus group. Annals of Applied Biology 113:493-505. Li, R.H. Zettler, FIW., Elliott, M.S., Petersen, M.A., Still, P.E., Baker, C.A. and Mink, G.I. 1991. A strain of peanut mottle virus seedborne in bambara groundnut. Plant Disease 7 5:130133. Linnemann, A.R. and Azam Ali, S. 1993.Bambara groundnut (Vigna subterranea). In: Pulses and Vegetables. Williams J.T. (Ed.), pp. 13-58. Chapman and Hall, London, U.K. Ng, N.Q., Thottapilly, G. and Gumedzoe, M. 1985. Virus Diseases. In: Annual Report for 1984. International Institute of Tropical Agriculture, Ibadan, Nigeria. 223pp. Purcifull, D.E. 1966. Some properties of tobacco etch virus and its alkaline degradation products. Virology 29:8-14. Rajeshwari, R., Lizuka, N., Nolt, B .L. and Reddy, D.V.R. 1983. Purification, serology and physico-chemical properties of a peanut mottle virus isolate from India. Plant pathology 32: 197-205. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Photo images[cs96045a.jpg] [cs96045b.jpg] |
| |||||||||

{kind=link}
{kind=link}