 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Genetic analysis of tassel size and leaf senescence and their relationships with yield in two tropical Lowland maize populations^1
R. G. GUEI and C. E. WASSOM^1 Warda 01 BP 2551 Bouake 01, Cote D'Ivoire ^1 Department of Agronomy, Kansas State University, Manhattan, Ks 66506-5501, USA (Received 16 February, 1996; accepted 16 August, 1996)
Code Number: CS96066
Sizes of Files:
Text: 19.5K
Graphics: Line drawings (gif) and Tables (jpg) - 315K
ABSTRACT
Primary tassel branch number, tassel weight, and leaf senescence are among plant traits which affect maize plant yielding efficiency. Two random mating tropical maize populations were used to estimate genetic components of variance and heritability, and to quantify the amount of genetic variability. Full-sibs within half-sib groups were developed using the mating Design I, and were evaluated at two locations for 2 seasons in Mexico. Results showed that large amounts of genetic variability were present in both populations for all traits. Additive genetic variance was greater in magnitude than dominance variance. Negative phenotypic and genotypic correlations were found between tassel size, leaf senescence, and yield. Selection based on these traits may be appropriate for increased yield. Overall, the inheritance of the traits measured depended on the genetic background of the population and the environmental conditions. Key Words: Additive variance, dominance variance, genetic variability, heritability, leaf senescence, tassel size RESUME Le nombre d'embranchements primaires de la fleur male, leur poids et la senescence des feuilles font parti des caracteristiques de la plante de mais qui affectent l'efficience du rendement. Deux populations de mais d'origine tropical croisees au hasard ont ete utilisees pour estimer les composantes de la variance genetique et de l'heritabilite, et pour quantifier l'ordre de grandeur de la variabilite genetique. Des groupes de vrais freres a l'interieur des demis freres ont ete developpes en utilisant le dispositif I de croisement, et ont ete evalues dans deux localites pendant deux saisons au Mexique. Les resultats demontrent que la variabilite genetique etait elevee dans les 2 populations pour toutes les caracteristiques mesurees. Mais la variance genetique additive etait plus elevee que la variance du type dominant. Les correlations phenotypiques et genotypiques etaient negatives entre les caracteristiques mesurees et le rendement. La selection basee sur ces caracteristiques pourrait ameliorer le rendement, et leur transmission serait fonction du determinisme genetique de la population et des conditions environnementales. Mots Cles: Heritabilite, Variabilite genetique, variance additive, variance dominante, senescence des feuilles INTRODUCTION A plant's efficiency can be measured by its ability to allocate most of the photosynthate produced toward the formation of grain. Traits such as plant height, ear height, leaf area, and leaf number can affect photosynthetic efficiency of maize (Zea mays L.) plants (Moss and Musgrave, 1971). Other important traits related to their efficiency are tassel size and leaf senescence of a plant, especially during grain filling. Tassel size affects grain yield, either physiologically by competition for photosynthates or physically by shading effect (Grogan, 1956; Hunter et al., 1969; Mock and Schuetz, 1974). Studies have shown that low yielding plants partition more photosynthate toward the formation of big and heavy tassels, instead of producing big ears (Hunter et al., 1969). Selecting for smaller tassel size should result in increased yield. Although tassel size, and leaf senescence are widely used as selection criteria in tropical maize, genetic studies are needed to provide breeders with information about the importance of gene action controlling the expression of these traits. Such information could aid plant breeders in the choice of breeding methods and resource allocation to achieve maximum gain. Therefore, the objectives of this study were (a) to estimate genetic components of variance and heritability of tassel size and leaf senescence and (b) to quantify the amount of genetic variability for these traits in two elite tropical maize populations.
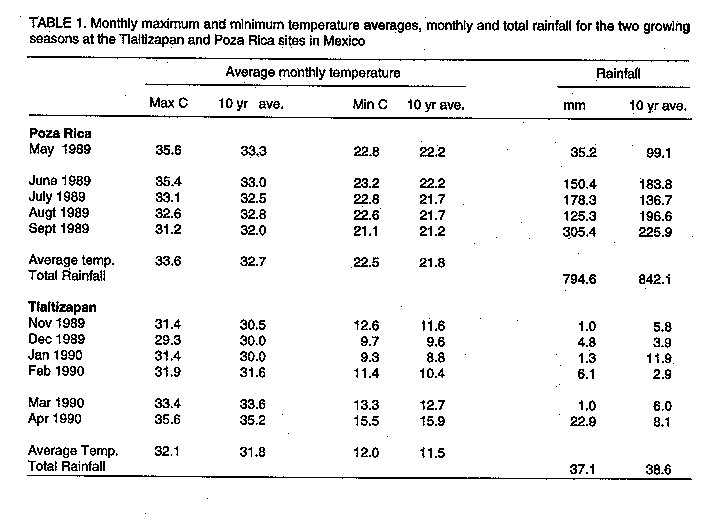
MATERIALS AND METHODS The study was conducted at the International Centre for Maize and Wheat Improvement (CIMMYT) in Mexico in 1989-1990. Two elite maize populations from CIMMYT (La Posta Sequia and Pool 26 Sequia) were used. La Posta Sequia is a late (120 days) maturity, lowland, tropical, white dent maize, often used in breeding programmes in West Africa for its resistance to maize streak virus. Pool 26 Sequia is a late (115 days) maturity, lowland, tropical, yellow flint/dent that is resistant to ear rots. Noninbred (S0) plants of La Posta Sequia and Pool 26 Sequia were planted in a breeding nursery at Poza Rica, Mexico in November 1988. Using the Design I mating system (Nested Design) introduced by Comstock and Robinson (1948), randomly selected males from each population (45 from La Posta Sequia and 42 from Pool 26 Sequia) were crossed to two randomly selected female plants. Progenies from La Posta Sequia (90 full-sib families) and Pool 26 Sequia (84 full-sib families) were evaluated at two locations for two seasons. The first evaluation was planted on May 15, 1989 at the Poza Rica CIMMYT station (latitude 200 32'N, longitude 97 degrees 26'W; elevation 60 m). Mean maximum and minimum temperatures during the growing season were 33.60 C and 22.50 C, respectively (Table 1). Rainfall during the test period was 794.6 mm which was slightly less than the 10 year average of 842.1 mm. The second trial, planted on November 15, 1989, was conducted in the dry winter season at Tlaltizapan, Morelos, Mexico (latitude 18degrees 41'N, longitude 99 degrees 08'W; elevation 940 m). Mean temperatures for the growing season were 32.10 C maximum and 120 C minimum. Rainfall during the test period at this site was 37.1 mm. A furrow irrigation system was used to apply 80 mm of water at both locations during the growing period as needed. The two-row plots (2.5 m long) with inter-row spacing of 75 cm were planted in two replications in Replication In Block Design (randomised incomplete-block design) after subdividing the males into sets or subgroups (9 sets each of 5 different males for La Posta and 7 sets each of 6 males for Pool 26). The sets were randomly allocated to blocks as were progeny rows within each set. Two seeds per hill were planted and seedlings were later thinned to obtain the required plant density (53,333 plants ha^-1). Sixteen competitive plants were chosen at random in each plot for data recording. Primary tassel branch number and tassel weight were determined about three weeks after flowering. Tassels were cut 1 cm below the first tassel branch and oven dried at 800 C for 4 days to uniform moisture content. Average primary tassel branch number and tassel weight per plot were obtained. About 2 weeks before harvest, each plot was scored on amount of green appearance (senescent or green). Scores were 1 for greenest plots to 5 for senescent plots. Scoring was done independently by three well trained people, and scores were averaged. Grain yield, number of ears per plant, days to 50% anthesis and silking and other data were presented earlier by Guei and Wassom (1992; 1993). Analyses of variance for plot averages for individual characters were performed as suggested by Hallauer and Miranda (1988). Variance components attributed to males (O^2[m]), females within males (O^2[f/m]) and their interactions with environments (O^2[em], O^2[ef/m]) were computed from the analysis of variance table (Table 2) using observed and expected mean squares. These components were used to estimate additive genetic variance (O^2[A]), dominance variance (O^2[D]), and their standard errors, ratio of dominance variance to additive variance, and narrow sense heritability, as estimated by Hallauer and Miranda (1988):
O^2 2[A] = 4 O^2[m] for noninbred plant (the inbreeding
coefficient F=0)
O^2 2D = 4 O^2 [t/m] - 4 O^2[m]
The standard errors of estimates of variance were computed by
taking the square root of the variance of the estimates. From
Table 2 the estimated variance of O^2[A] is:
Where e = number of environments
r = number of replications
O^2[c] = error varianceIn this study we assumed a random mating population and the absence of epistasis. Genetic coefficient of variation (cvg) was calculated as (Square Root of V[g] / x ) x 100, where V[g] is the genetic variance, and x is the mean of a trait (Kang et al., 1990). Genotypic and phenotypic correlations were computed according to Robinson et al. (1951) as follows:
Where ^rP[xy] = phenotypic correlation between
trait x and trait y
^g[xy] = genotypic correlation between trait x and
trait y
COVP[xy] = phenotypic covariance between trait x and
trait y
COVg[xy] = genotypic covariance between trait x and
trait y
O^2p[px] = phenotypic variance of trait x
O^2[py] = phenotypic variance of trait y
O^2[gx] = genotypic variance of trait x
O^2[gy] = genotypic variance of trait yData from each sample of progenies in each population were analysed. The form of the analysis is given in Table 2. RESULTS AND DISCUSSION
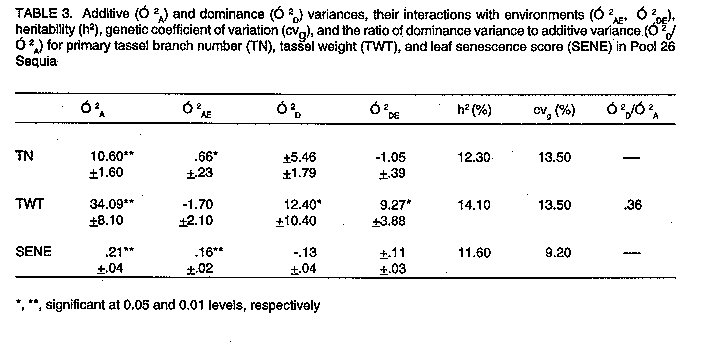
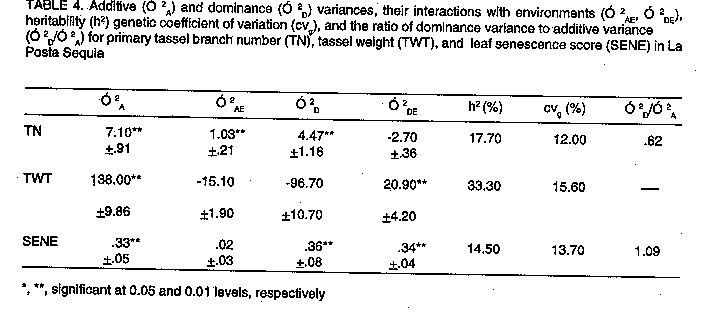
Pool 26 Sequia. Components of additive (O^2[A]) and dominance (O^2[D]) variances, their interactions with environments (O^2[AE], O^2[DE]), heritability (h^2), genetic coefficient of variation (cv[g]), and the ratio of dominance variance to additive variance (O^2[D]/O^2[A]) are given in Table 3 for primary tassel branch number (TN), tassel weight (TWT), and leaf senescence score (SENE). Additive genetic effects were significant (P < 0.01) in the inheritance of tassel characteristics and leaf senescence score. Mock and Schuetz (1974) found additive gene action to be important in the inheritance of tassel branch number. They also reported that the non-additive gene action was mainly dominance, and no epistasis was present. However, our results showed significant dominance gene action (P < 0.05) only in the expression of tassel weight. The relative importance of additive genetic variance to dominance indicated that fast progress from selection may be achieved for tassel size and leaf senescence score in this population. The expression and magnitude of the additive genetic variance depended upon the environmental conditions as the environment by additive variance interactions for TN (P < 0.05), and SENE (P < 0.01) were significant. The interaction (O^2[AE]) for TWT was not significant. On the other hand, O^2[DE] was not significant for TN and SENE, but was significant for TWT. The heritability estimate, although not large, was higher for TWT (14.1) than for TN (12.3) and SENE (11.6). High heritability estimates have been reported for TN in previous works (Mock and Schuetz, 1974; Geraldi et al., 1985), and relatively low estimates may have resulted from limited testing, i.e., higher heritability estimates might have been obtained if more environments had been included in this study. Large estimates of heritability for TN, TWT and SENE would justify the use of visual selection often used to differentiate among genotypes for tassel size and/or green vs senescent plots. In such a case, visually scoring plots based on tassel size would provide an easy alternative to counting primary tassel branches as was done in this study. Large genetic variability was present in this population for these traits, as shown by the genetic coefficients of variation (cvg) for TN (13.5) and TWT (13.5). Cvg was somewhat lower for SENE (9.2). The ratio O^2[D]/O^2[A] for TWT (0.36) showed the importance of the additive genetic variance compared to dominance, and indicated that the primary gene action may be partial to complete dominance (Robinson et al.,1960). When the dominance variance was negative, variance ratios were not estimated. Negative phenotypic and genotypic correlations with yield were obtained for all traits, but were only significant (P < 0.05) for TWT and SENE (Table 5). La Posta Sequia. In this population, both additive and dominance variance were important in controlling the expression of TN and SENE (Table 4). These results differed from the findings in Pool 26 Sequia where only additive gene action was found to be important in the expression of these traits. Similar results were reported by Mock and Schuetz (1974) who showed the importance of additive and dominant gene effects in the inheritance of TN, although the majority of genetic variation was due to additive gene action. The ratio O^2D/O^2[A] (0.62) less than one, confirmed the effectiveness of additive genetic variance in controlling the expression of TN. However, O^2[D]/O^2[A] (1.09) for SENE, showed that dominance gene effects were predominant in this trait, contrary to TN. O^2[D]/O^2[A] for these two traits were greater than 0.50, suggesting that overdominance could be the mode of expression of genes controlling these traits (Robinson et al., 1960). The interactions between additive genetic variance and environmental variance were not significant for tassel weight and senescence score; however, the dominance by environment interactions were significant (P < 0.01). Large genetic coefficients of variation obtained for all traits demonstrated that large amounts of genetic variability were present. Somewhat more variability was present in TWT (cvg = 15.6) than TN (12) and SENE (13.7). The heritability estimate for TWT was 33.3 as compared to 14.1 for Pool 26 Sequia. These results indicated that mass selection for tassel size might be more efficient in La Posta Sequia than Pool 26 Sequia. Phenotypic and genotypic correlations with yield were negative for all traits, indicating that the lower the tassel size and leaf senescence score, the higher the yield (Table 5). SUMMARY
The mating Design I was used in this study to characterize genetic variation due to additive and nonadditive effects in two random mating tropical lowland maize populations. These elite populations are being developed at CIMMYT, Mexico for drought tolerance. Results showed that large amounts of genetic variability existed for primary tassel branch number, tassel weight and leaf senescence. Additive genetic effects were more important than dominance effects in controlling the expression of the traits in both populations. Dominance variance was significant (P < 0.01) for TN and SENE in La Posta Sequia but not in Pool 26 Sequia. La Posta Sequia has had a longer history of selection than Pool 26 Sequia because it was derived from population Tuxpe–o, which has been carried through eight selection cycles for drought. This may have contributed to the buildup of dominance variance (Hull, 1945). For selection purposes, additive genetic variance is of primary importance and thus progress should be expected from selection for increased yield. Greater heritability estimates were obtained in La Posta Sequia, suggesting that mass selection for tassel size might be more efficient in this population than in Pool 26 Sequia. Negative correlations indicated that selecting for smaller tassel size, and greener plants at maturity may increase yield. Overall, the inheritance of tassel size characteristics and leaf senescence depended on the genetic background of the population and the environmental conditions. ACKNOWLEDGEMENTS
This paper is part of a dissertation submitted by the first author in partial fulfillment of the requirements for the Ph.D. degree, Department of Agronomy, Kansas State Univesity, Manhattan, Ks 66506-5501. We are grateful to the CIMMYT maize programme and to Pioneer Hi-Bred Seed Company, Ames, Iowa USA for their support. REFERENCES Comstock, R.E. and Robinson, H.F. 1948. The components of genetic variances in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4:254-266. Geraldi, I.O., Miranda Filho,J.B. and Vencovsky, R. 1985. Estimates of genetic parameters for tassel characters in maize (Zea mays L.) and breeding perspectives. Maydica 30:1-14. Guei, R.G. and Wassom, C.E. 1992. Inheritance of some drought adaptive traits in maize: I.Interrelationships between yield, flowering, and ears per plant. Maydica 37:157-164. Guei, R.G.and Wassom, C.E. 1993. Genetics of osmotic adjustment in breeding maize for drought tolerance. Heredity 71:436-441. Grogan, C.O. 1956. Detasseling responses in corn. Agronomy Journal 48:247-249. Hallauer, A.R. and Miranda, J.B. 1988. Heredity variance: Mating designs. In: Quantitative Genetics in Maize Breeding, pp. 45-114. Iowa State University Press/Ames. Hull, F.H. 1945. Recurrent selection for specific combining ability in corn. Journal of American Society of Agronomy 37:134-145. Hunter, R.B., Daynard, T.B., Hume, D.J., Tanner, J.W., Curtis,J.D. and Kannenberg, L.W. 1969. Effect of tassel removal on grain yield of corn. Crop Science 9:405-406. Kang, M.S., Sosa, D. and Miller, J.D. 1990. Genetic variation and advance for rind hardness, flowering, and sugar yield traits in sugarcane. Field Crops Research 23:63-73. Mock, J.J. and Buren, L.L. 1972. Classification of maize (Zea mays L.) inbreds for population tolerance by general combining ability. Iowa State Journal of Science 46:395-404. Mock, J.J. and Schuetz, S.H. 1974. Inheritance of tassel branch number in maize. Crop Science 14:885-888. Moss, D.N. and Musgrave, R.B. 1971. Photosynthesis and crop production. Advance in Agronomy 24:317-334. Robinson, H.F., Cockerham, C.C. and Moll, R.H. 1960. Studies on estimation of dominance variance and effects of linkage bias. In: International Series of Monographs on Biometry. Kempthorne, O. (Ed.), pp. 171-177. Proceedings of an International Symposium of Ottawa, August 1958. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Photo images[cs96066g.jpg] [cs96066c.jpg] [cs96066d.jpg] [cs96066f.jpg] [cs96066e.jpg]Line drawing images[cs96066a.gif] [cs96066b.gif] |
| |||||||||

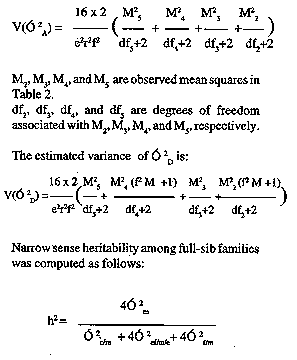{kind=link}
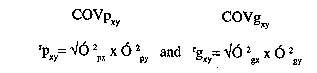{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}