 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol.5. No.2, pp. 1997 Inheritance of leaf mark, pod dehiscence and dry pod colour in crosses between wild and cultivated cowpeas V. O. ALIBOH, O.B. KEHINDE and I. FAWOLE* Department of Agricultural Biology, University of Ibadan, Ibadan, Nigeria. (Received 5 July, 1995; accepted 18 February, 1996)
Code Number: CS96067
Sizes of Files:
Text: 19.8K
Graphics: Tables (jpg) - 173.5K
ABSTRACT
The inheritance of three traits, inverted V-shaped mark on leaves, pod dehiscence and dry pod colour, was studied in crosses between wild, weedy and cultivated varieties of cowpea, Vigna unguiculata (L) Walp. By cross-pollinating only open flowers that failed to produce pollen, successful crosses were obtained on wild cowpea as female parents in contrast to previous attempts. Inheritance data collected on the parents, Fl, F2 and backcross progenies, indicate that each of the three traits is controlled by a single dominant gene in the crosses that were examined. The wild and the weedy lines carry dominant genes while the recessive alleles reside in the cultivated varieties but the genes controlling the traits, in both wild and weedy cowpea lines are allelic. The symbols Vsm, Dhp and Bk-2 are assigned to the dominant genes that govern the presence of the inverted V-shaped mark on leaves, pod dehiscence and black dry pod colour, respectively. The significance of these results in cowpea improvement is discussed. Key Words: Cowpea genetics, Vigna unguiculata, dominant genes RESUME
L'heredite des trois caracteres, la forme V inversee sur les feuilles, dehiscence des gousses, la couleur des gousses sechees a ete etudiee ˆ la suite des croisements entre les varietes sauvages herbacees et cultivees du niebe Vigna unguiculata (L) Walp. En croisant seulement des fleurs ouvertes pendant la pollinisation qui n'ont pas produit des pollens, des parents femelles ont ete obtenus avec le niebe sauvage suite aux croisements reussis contrairement aux precedentes tentatives. Les donnees hereditaires collectees sur les parents, F1 F2 et les descendants du "Back cross" montrent que chacun des trois caracteres est controlle par un seul gene dominant sur les croisements examines. Les lignees sauvages et herbacees portent des genes dominants alors que les alleles recessifs resident dans les varietes cultivees mais les genes controllant les caracteres dans les deux lignees du niebe (Sauvage et herbacee) sont sur les alleles. Les symboles Vsm, Dhp et Bk-2 sont assignes sur les genes dominants qui gouvernent respectivement la presence de la form V inversee sur les feuilles, la dehiscence des gousses, la couleur des gousses sechees. L'importance de ces resultats sur I' amelioration du niebe est evoquee. Mots Cles: Vigna unguiculata, la genetique du niebe, genes dominants
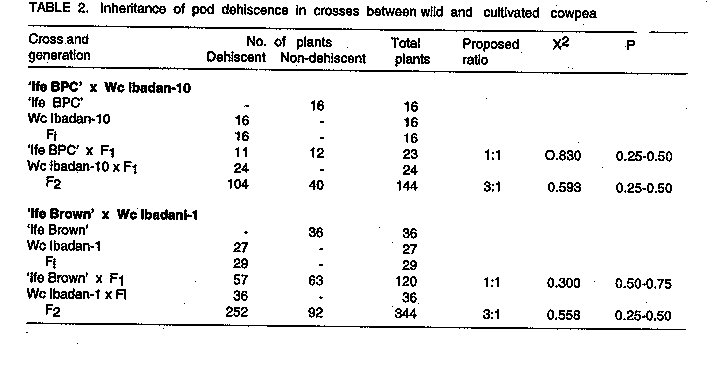
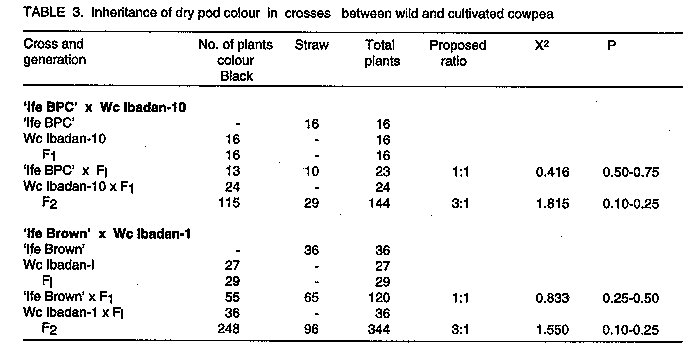
INTRODUCTION Cowpea, Vigna unguiculata (L.) Walp., is a widely cultivated crop in the tropical and subtropical regions of the world. Cowpea is native to West Africa and wild and weedy forms exist in many parts of the region (Faris, 1965; Rawal, 1975; Ng and Marechal, 1985). Wild relatives of crop species are often sources of genes for disease and insect resistance, increased yield, improved product quality, earliness and wide adaptation (Leppik, 1970; Stalker, 1980). Wild forms and closely related species of cowpea, therefore, have great potential as an additional source of useful germplasm for cowpea improvement (Baudoin and Marechal, 1985; Padulosi and Ng, 1990). The wild and the weedy subspecies of cowpea (V. unguiculata subsp. dekintiana, stenophylla etc.) hybridise easily with the cultivated forms and produce viable hybrids (Rawal, 1975; Evans, 1976; Ng and Marechal, 1985; Baudoin and Marechal, 1985; Ng, 1990). But according to Rawal et al. (1976), the wild form could only be used as the male parent and attempts to use it as the female parent were unsuccessful. In order to utilise wild relatives of cowpea effectively for cultivar improvement, the inheritance patterns of both useful and undesirable traits in the wild relatives must be known. The objective of this study, therefore, was to determine the inheritance of three traits, the inverted V-shaped mark on leaves, pod dehiscence, and dry pod colour in crosses between cultivated cowpea and wild varieties belonging to the subspecies dekindtiana. MATERIALS AND METHODS Eight cowpea lines, consisting of five cultivated, two wild and one weedy varieties, were used in the study. The cultivated varieties are TVu 1, TVu 4552, TVu 6198, 'Ife Brown' and 'Ife BPC'. Their morphological characteristics are described by Porter et al. (1974) and Fawole and Afolabi (1983). TVu 1 pods are wholly pigmented and the pigmentation is retained in the dry pods. The other varieties have straw coloured, non-dehiscent dry pods. All the varieties are devoid of the inverted V-shaped marks on their leaves. The wild and the weedy lines were taken from the collection maintained in the Department of Agricultural Biology, University of Ibadan, Nigeria. The lines were originally collected around Ibadan (latitude 7o 30' N) in habitats similar to those described by Rawal (1975). The wild lines, designated Wc Ibadan-1 and Wc Ibadan-8 have a climbing growth habit. The leaves are subhastate in shape with very distinct, white, inverted V-shaped marks connecting both margins in each of the three leaflets. Flowers are large and aromatic while dry pods are black in colour and shatter with considerable force, scattering the small, smooth self-coloured seeds. The weedy line, Wc Ibadan-10, is an annual indeterminate plant. The leaves are subglobose in shape and possess the inverted V-shaped mark on the leaflets. The flowers are purple-coloured and are comparable in size to those of the cultivated varieties but pod characteristics are similar to those of the wild form. Seeds are small, smooth and solid tan in colour. According to a 1978 classification by Marechal and his associates (cited in Marechal and Ng, 1985), both of these forms belong to the subspecies dekindtiana of Vigna unguiculata (L.) Walp. The wild form was referred to as the variety mensensis of this subspecies while the weedy form was classified as the variety dekindtiana. Seeds of the wild lines were planted in plastic pots in a glass-house in August of 1985 and those of the other lines except 'Ife Brown' were planted a month later to synchronise flowering with the wild lines. Seeds of the wild and weedy lines were scarified before planting to promote early and uniform germination. At the onset of flowering, reciprocal crosses were made between the lines using the method described by Rachie et al. (1975). This method did not produce successful crosses when the wild lines were used as female parents. To obtain crosses on the wild lines as female parents, open flowers of the lines were examined early in the morning for the presence or absence of pollen. Flowers that did not produce pollen were assumed to be male sterile and were cross-pollinated. Seeds from individual pods resulting from such cross pollinations were planted separately to ascertain whether the crosses were successful. Backcross seeds were also produced for some selected crosses. In order to investigate whether cytoplasmic factors affect the inheritance of the traits, the following selected crosses and their reciprocals were studied; 'Ife BPC' x Wc Ibadan-1, TVu 1 x Wc Ibadan-1, TVu 4552 x Wc Ibadan-1, TVu 6198 x Wc Ibadan-1, 'Ife BPC' x Wc Ibadan-8, 'Ife BPC' x Wc Ibadan-10, TVu 1 x Wc Ibadan-10, and TVu 4552 x Wc Ibadan-10. The sixteen crosses and their seven parental lines were grown in a randomised complete block experiment with four replicates on the roof top of the Department of Agricultural Biology, University of Ibadan in September of 1986. A plot was a 25 cm plastic pot filled with sterilised garden soil. Daily watering of the plants was carried out and hand weeding was done as necessary. Monocrotophos was used to control insect pests. Three weeks after planting, and at the onset of flowering, individual plants were scored for the presence or absence of the inverted V- shaped mark on the leaves. At maturity, the pods of individual plants were classified as dehiscent or indehiscent and pod colour was categorised as black, or straw coloured. An inheritance study of the traits was carried out on three crosses, TVu 1 x Wc Ibadan-1, 'Ife BPC' x Wc Ibadan-8 and ÔIfe BPC' x Wc Ibadan-10, grown in pots on the roof top garden of the Department of Agricultural Biology, University of Ibadan in November of 1987. Each cross consisted of six genetic families, namely, Pl, P2 and Fl, BCP1, BCP2 and F2. The design used for the experiment, was the randomised complete block with four replicates. A replicate consisted of 60 pots: Pl, P2 and Fl were randomly assigned to four pots each, the two backcrosses were assigned to six pots each while the remaining 36 pots were grown to the F2, giving a total of 240 plants for each cross. Data were taken on the traits as indicated earlier. In the cross 'Ife BPC' x Wc Ibadan-8, several F2 plants, especially those with the branched peduncle trait, were extremely late flowering and could not be scored for pod characteristics. Similarly, the cross TVu 1 x Wc Ibadan-1 was not scored for the pod traits because TVu 1 was not completely indehiscent under the experimental conditions and the presence of anthocyanin pigment on the pods did not permit classification of dry pod colour into distinct phenotypic classes. Two additional crosses, 'Ife Brown' x Wc lbadan-1 and Wc Ibadan-10 x Wc Ibadan-1 were made in March of 1990. During the second planting season of 1990, the crosses were planted on the Teaching and Research Farm on rows six metres long with a spacing of 90 cm between rows and 30 cm within rows. The cross ÔIfe Brown' x Wc Ibadan-1 consisted of Pl, P2, Fl, F2 and the two backcrosses while Wc Ibadan-10 x Wc Ibadan-1 consisted only of Pl, P2 Fl and F2 generations. Hand weeding of the plots was done and insect pests were controlled as necessary. Data on individual plants were taken as described for the other crosses. The data obtained in all the experiments were analysed and the segregation data were tested for their goodness-of-fit to the appropriate genetic ratios by the Chi-square test. RESULTS Reciprocal F1, crosses between wild, weedy and cultivated varieties were successful, thus permitting a test for the presence of cytoplasmic factors in the control of the traits under study. All the leaves of reciprocal Fl plants had the inverted V-shaped mark. Similarly, the dry pods of reciprocal Fl plants were dehiscent and black in colour. These results indicate the absence of maternal or cytoplasmic factors in the control of these traits. Data on the inheritance of the V-shaped mark on leaves, pod dehiscence, and pod colour are shown in Tables 1-3. In all cases, the leaves of Fl plants and those of the backcross to the wild and the weedy parents had the inverted V-shaped mark (Table 1) while the dry pods were dehiscent (Table 2) and black in colour (Table 3), indicating that these traits, which occur in the wild and the weedy lines, were completely dominant over the normal types found in the cultivated varieties. Progenies of the backcross to the cultivated parents segregated in a 1:1 ratio while in the F2, segregation ratio was 3:1 for each pair of genes. These results suggest that the presence of the inverted V-shaped mark on leaves, dry pod dehiscence, and black dry pod colour are each controlled by a single dominant gene. Allelic tests of the dominant genes in the wild and the weedy lines were carried out. If allelic, the Fl and F2 progenies of the cross Wc Ibadan-1 x Wc Ibadan-10 should have the wild-type phenotype and if non-allelic, the F2 progeny should segregate in the ratio 15 wild-type:l alternative phenotype. All the 32 Fl and 291 F2 plants of the cross had the inverted V-shaped mark on their leaves while the dry pods were dehiscent and black in colour. Thus the inverted V-shaped mark on leaves, pod dehiscence and black dry pod colour were each under the control of the same genes in both wild and weedy lines. The symbol Vsm is assigned to the dominant gene that governs the presence of the inverted V-shaped mark on leaves while the symbol Dhp is assigned to the dominant gene for pod dehiscence. Capinpin (cited by Fery, 1985) described a dominant gene controlling black pod colour in a varietal cross of cowpea and suggested the symbol B for the gone while Fery (1985) redesignated the gene Bk. Since the line carrying the dominant gene in Capinpin's cross was not available for allelic test, the symbol Bk-2 is proposed for the black dry pod colour gene in the wild and the weedy lines used in this study.
DISCUSSION Crosses using the wild varieties as female parents were successful when open flowers that failed to produce pollen were cross-pollinated but not when the standard method of emasculation and pollination was employed. It appears, therefore, that there are no physiological barriers to the successful use of wild cowpea as female parents in crosses. Rawal et al.(1976) reported unsuccessful attempts to use wild forms of cowpea as female parents in crosses. This might be due to the high degree of sensitivity of the flowers to disturbance in the process of emasculation. The successful crosses were obtained during the dry season when relative humidity was low, an environmental condition that seemed to favour non-production of pollen in some flowers of otherwise fertile plants. The absence of reciprocal differences in the crosses suggests that either the wild and the weedy lines or the cultivated varieties may be used as female parents. In practice, however, it may be more convenient to use cultivated varieties as female parents because of the greater probability of obtaining successful crosses. In the wild and the weedy cowpeas, the inverted V-shaped mark is clearly visible at the first true leaf stage of plant development. If the trait is introduced from the pollen, it may be used as a marker to identify successful crosses at the seedling stage thus saving time and resources. In addition, it can be used to establish linkage relationships and if found to be linked with mature plant characters, it can be an aid to selection. Pod shattering or dehiscence is a seed dispersal mechanism that enhances the survival of the plant in the wild. However, it is an undesirable trait in cultivated varieties. Since the trait is controlled by a single dominant gene, it should not be difficult to eliminate it from breeding populations. The role of the black dry pod colour in the evolution of wild cowpea is not known. General observations in segregating populations of crosses between wild and cultivated cowpeas suggest that pod colour has no deleterious effects on agronomic traits but whether it confers any special advantage is yet to be determined.
ACKNOWLEDGEMENT
The authors thank the University of Ibadan Senate Research Grants Committee for financial support. REFERENCES Baudoin, J.P. and Marechal, R. 1985. Genetic diversity in Vigna. In: Cowpea Research, Production and Utilization. Singh, S.R. and Rachie, K.O. (Eds.), pp. 3 - 9. Chichester, John Wiley and Sons. Evans, A.M. 1976. Species hybridization in the genus Vigna. In: Proceedings of a Collaborators' Meeting on Grain Legume Improvement, 9 - 13, June 1975. Pages 31 - 34. International Institute of Tropical Agriculture, Ibadan, Nigeria. Faris, D.G. 1965. The origin and-evolution of the cultivated forms of Vigna sinensis. Canadian Journal of Genetics and Cytology 7:433-452. Fawole, I. and Afolabi, N.O. 1983. Genetic control of a branching peduncle mutant of cowpea, Vigna unguiculata (L.) Walp. Journal of Agricultural Science (Cambridge) 100:473-475. Fery, R.L. 1985. The genetics of cowpeas: a review of the world literature. In: Cowpea Research, Production and Utilization. Singh, S.R. and Rachie, K.O. (Eds.), pp. 25 - 62. Chichester, John Wiley and Sons. Leppik, E.E. 1970. Gene centres of plants as sources of disease resistance. Annual Review of Phytopathology 8:323-344. Ng, N.Q. and Marechal, R. 1985. Cowpea taxonomy, origin and germplasm. In: Cowpea Genetic Resources. Singh, S.E. and Bachie, K.O. (Eds.), pp. 11 - 21. International Institute of Tropical Agriculture, Ibadan, Nigeria. Ng, N.Q. 1990. Recent development in cowpea germplasm collection, conservation, evaluation and research at the Genetic Resources Unit, I.I.T.A. In: Cowpea Genetic Resources. Ng, N.Q. and Monti, L.M. (Eds.), pp. 13-28. International Institute of Tropical Agriculture, Ibadan, Nigeria. Padulosi, S. and Ng, N.Q. 1990. Wild Vigna species in Africa. Their collection and potential utilization. In: Cowpea Genetic Resources. N.Q. Ng and L.M. Monti (Eds.) pp. 58-77. International Institute of Tropical Agriculture, Ibadan, Nigeria. Porter, W.M., Rawal, K.M., Rachie, K.O., Singh, S.R., Wien, H.C. and Williams, R.J. 1974. The Cowpea Germplasm Catologe No. 1. International Institute of Tropical Agriculture, Ibadan, Nigeria. Rachie, K.O., Rawal, K.M. and Franckowiak, J.D. 1975. Rapid hand crossing of cowpea. International Institute of Tropical Agriculture Technical Bulletin 2. Ibadan, Nigeria. Rawal, K.M. 1975. Natural hybridization among wild, weedy and cultivated Vigna unguiculata (L.) Walp. Euphytica 24:699-707. Rawal, K.M., Rachie, K.O. and Franckowiak, J.D. 1976. Reduction in seed size in crosses between wild and cultivated cowpeas. Journal of Heredity 67:253-254. Stalker, H.T. 1980. Utilization of wild species for crop improvement. Advances in Agronomy 33:111-147. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Photo images[cs96067a.jpg] [cs96067c.jpg] [cs96067b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}