 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Interaction between soybean mosaic potyvirus and Meloidogyne incognita infection C.C. IHEUKWUMERE, G. I. ATIRI^1, B. FAWOLE^1 and K.E. DASHIELL^2
Department of Biological Sciences, University of Agriculture,
Markudi, Nigeria. (Received 8 January, 1996; accepted 29 August, 1996)
Code Number: CS96075
Sizes of Files:
Text: 22.7K
Graphics: Line drawing (gif) - 17K
Table (jpg) - 95K ABSTRACT Infection of soybean (Glycine max (L.) Merrill) by soybean mosaic potyvirus (SMV) and root-knot nematode, Meloidogyne incognita (Kofoid and White) Chitwood, occurs wherever soybean is grown in Nigeria. The effect of concomitant infections of four soybean cultivars by SMV (isolate 10) and root-knot nematode (race 2) was investigated in greenhouse, experimental field and on-farm studies. Soybean mosaic potyvirus was introduced into soybean test plants by mechanical inoculation of homogenised crude extract from infected leaves suspension into a shallow trench around the base of test plants. In all four soybean cultivars, simultaneous or successive inoculation with both the nematode and the virus resulted in more severe symptoms and, therefore, reduced yield and yield components when compared with inoculations with each pathogen alone, although the difference was not much with plants inoculated with virus only. The results were more variable under experimental field and on-farm conditions than under more uniform greenhouse conditions. However, there was a significant interaction (P=0.05) between inoculation treatments and cultivars. Since multi-pathogen infections depressed yields more than single infections, more emphasis should be placed on evaluating or breeding for combined resistance to both pathogens in soybeans. Key Words: Glycine max, multi-pathogen infections, root-knot nematode, virus RESUME L'infection du soja (Glycine max (l.) Merril) par le virus de la mosaque du soja (soybean mosaic potyvirus (SMV) et du nematode des racines, Meloidogyne incognita (Kofoid et white) Chitwood, a cours partout ou le soja est cultive au Nigeria. Let des infections de quatre cultivars du soja par SMV (isolate 10) et de nematodes des racines (race 2) etait introduit dans les plantes de soja par inoculation mecanique draits bruts homognes de feulles infestees, alors que lculation du nematode des racines etait realisee en versant une suspension dfs du nematode dans une tranchee peu profonde autour de la base des plant test. Pour les quatre cultivars de soja, lculation simultanee ou successive avec la fois le nematode et le virus a donne lieu des synptomes plus sevres et a par consequent entraine une baissse de rendement separement comparee aux inoculations des agents pathognes, bien que la difference nit pas sensible avec les plantes inoculees avec les seuls virus. Ces resultats entre cultivars etaient plus variables en champs experimentaux et en milieu reel que dans le serre ou les conditions sont plus uniformes. Cependant, il y avait une interaction (P=0.05) sensible entres les traitement dculation et de cultivars. Ds lors que lection avec une multitude dnts pathognes a occasione une baisse de rendement superieure a celle avec un seul pathogne, lent devrait tre mis sur lluation ou la selection en vue d resistance combinee deux agents pathognes du soja. Mots Cles: Glycine max, infections multi-pathognes, nematode, virus INTRODUCTION Soybean mosaic potyvirus (SMV) is a common pathogen wherever soybean (Glycine max (L.) Merril) is grown in Nigeria (Rossel and Thottappily, 1985; Thottappilly, 1992). The root-knot nematode (RKN), Meloidogyne incognita, is also widely distributed in these production areas (Adesiyan et al., 1990). Each may severely limit the yield of soybean (Riggs and Schmitt, 1987; Chen et al., 1994), a crop that is becoming increasingly important in Nigeria as a protein-rich source (Anon., 1990). Although RKN has not been implicated as a vector of plant viruses, it is known to cause more severe effects in multi-pathogen infections with some viruses (Adesiyan et al., 1990). The effect of multi-pathogen infection may depend on whether both virus and nematode occur simultaneously or successively. For instance, growth of tomato (Lycopersicon esculentum Mill) plant was significantly reduced in simultaneous inoculations with RKN and tobacco mosaic virus, but not when the virus preceded nematode inoculation (Goswani and Chenulu, 1974). A similar effect was also noted in maize mosaic virus and RKN interaction in maize (Zea mays L.) (Khurana et al., 1970). In preliminary survey, mixed infection with SMV and RKN have been frequently observed in major soybean growing areas in Nigeria. In this study, the effect of concomitant infections of SMV and RKN were investigated in four soybean varieties with a range of reaction to these pathogens.
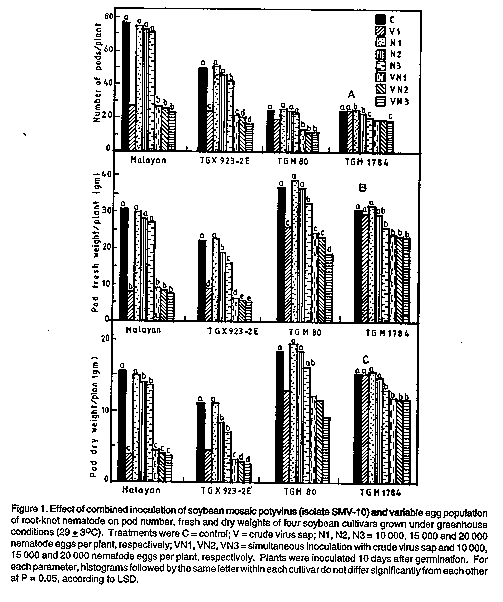
MATERIALS AND METHODS Source and maintenance of inocula. A widely distributed strain of SMV (isolate SMV-10) in the major soybean growing areas of Nigeria was provided by Dr. Thottappily (International Institute of Tropical Agriculture, IITA, Ibadan). It was propagated and maintained by collecting infected soybean leaf samples. These were homogenised in 0.01M pH 7.0 phosphate buffer in the ratio of 1:10 (w/v), and used in mechanically inoculating carborundum (600 mesh)-dusted leaves of 2-wk-old "Malayan" soybean seedlings in the greenhouse (29 +/- 3 C), according to standar procedures (Walkey, 1991). The most prevalent race of RKN in Western Nigeria, M. incognita race 2, was obtained from roots of Celosia agrgentea L., a highly susceptible host plant. The identity was confirmed using perineal patterns as described by Eisenback et al., (1981). The nematode was multiplied by inoculating roots of 3-week-old Celosia plants, from which eggs were extracted after 8 weeks (Hussey and Barker, 1973). Numbers of eggs neede for inoculation were calculated according to the method of Hartman and Sasser (1985). Greenhouse test. The four soybean cultivars used in these tests were malayan (highly susceptible to SMV, but moderately resistant to RKN). "TGX 923-2E" (Susceptible to both), "TGM 80" (moderately resistant to SMV but susceptible to RKN) aand "TGM 1784" (resistant to both) (Iheukwumere et al., 1995). In May 1991, five seeds of each cultivar were sown in 15-cm-diameter plastic pots containing 2.5 kg sterile sandy-loam soil. Seedlings were later selected for uniformity by thinning to one plant per plot one week after emergence. The following treatments were applied 10 days after germination: (i) mechanical inoculation of primary leaves with crude virus sap (V), (ii) inoculation of the roots with 10,000 nematode eggs per plant (N1), iii) inoculation of the roots with 15,000 nematode eggs per plant (N2), iv) inoculation of the roots with 20,000 nematode eggs per plant (N3), v) simultaneous inoculation with crude virus sap and 10,000 nematode eggs per plant (VN1), vi) simultaneous inoculation with crude virus sap and 15,000 nematode eggs per plant (VN2), vii) simultaneous inoculation with crude virus sap and 20,000 nematode eggs per plant (VN3), viii) uninoculated plants. Mechanical inoculation with SMV was carried out as described above. To inoculate RKN, the egg suspension containing the required number of eggs was poured into a shallow trench around the base of the plant and covered immediately with top soil (Goswani and Chenulu, 1974). The levels of nematode egg inocula are those that would normally initiate root-knot infection in a susceptible plant. The plants were completely randomised on greenhouse benches maintained at (29 +/- 3 C). Plants were spaced 50 x 50 cm and were watered every other day by sprinkler to reduce soil splash and cross contamination. Plants were inspected weekly for symptom development. The experiment was terminated when the plants were fully mature and 95% of the pods had attained mature pod colour (Feh and Caviness, 1977), about 100 days after sowing. At that time, the number and fresh and dry weight of pods were measured. Field and on-farm trials. These were carried out from April to August, 1992, at the IITA, Ibadan, and on-farm location at Ayepe, the latter being part of the IITA Resource Crop Management Programme. The two areas are about 50 km apart, have a bimodal rainfall pattern (i.e., with two peak periods of rainfall annually), with soils that are predominantly alfisols and entisols (Mutsaers, 1991). In both locations, the experimental design was a split plot consisting of four blocks (17.5 x 3.6 m each), each of which was divided into four mainplots assigned to one of four cultivars described in the greenhouse test above. Each mainplot was further subdivided into five subplots (2.4 x 2.3 m each) in which the following treatments were randomised: i) plants inoculated with only nematodes (20, 000 eggs per plant) 10 days after germination (N), ii) plants inoculated with the nematodes followed by the virus 7 days later (N + V7), iii) plants inoculated with the nematodes followed by the virus 14 days later (N + V14), iv) plants inoculated with virus 10 days after germination (V), and v) inoculated plants (C). The SMV and RKN inoculation methods were as described for the greenhouse test above. There were five rows per plot and plants were spaced 5 cm along rows that were 60 cm apart. To prevent movement of RKN from plots with plants inoculated with RKN to plots inoculated with SRM and RKN or plants inoculated with SMV alone, the granular nematicide carborfuran (as Furadan, 10% a.i., FMC, New York) was incorporated into the soil in plots with V and C treatments at the rate of 10 kg ha^-1 prior to planting. When pants were mature (about 100 days after sowing), 18 plants were harvested from the middle rows of each plot, while the first and fifth rows were not sampled to reduce border effects. Seed yield in kg ha^-1 at 130 g^-1 moisture content was recorded, means were calculated, and an analysis of variance (ANOVA) performed. The Least Significant Difference (LSD) was determined where significant differences were detected (Gomez and Gomez, 1984). RESULTS Greenhouse studies. Effect of combined inoculation of virus and variable nematode inoculum levels on soybean pod number, fresh and dry weight was evaluated in a greenhouse study in Malayan. Plants inoculated either with virus alone or simultaneously with the nematode did not differ significantly in numbers of pods per plant, although pod numbers were significantly reduced relative to the inoculated control or plants inoculated with the nematode alone (Fig. 1A). A similar trend was observed for pod fresh and dry weights (Fig. 1B and 1C). In TGX 923-2E, there were no significant differences in pod numbers among inoculated plants and plants inoculated with either 10,000 or 15,000 nematode eggs (Fig. 1A). However, inoculation with the virus alone, nematodes (20, 000 eggs), and all combinations of virus and nematode significantly reduced pod numbers compared to uninoculated plant. Multi-pathogen infections reduced pod numbers more than single infections with the nematode. Pod fresh and dry weights were also significantly reduced relative to the control in plants inoculated with the virus alone, nematode alone (at 15,000 and 20,000 eggs) and all combinations of the virus and nematode (Fig. 1B and 1C). Multi-pathogen infections with SMV and RKN at 20 000 eggs reduced these parameters more than single infections. Multi-pathogen infections by SMV and RKN at all inoculum levels significantly reduced pod numbers in TGM 80, while all other treatments did not differ significantly from uninoculated plants (Fig. 1A). Only plants inoculated with RKN alone at 10,000 and 15,000 eggs did not differ significantly from uninoculated control plants in pod fresh and dry weight (Fig. 1B and 1C). Two distinct trends emerged in TGM 1784. Plants inoculated with SMV or RKN alone at 10, 000 and 15,000 eggs did not differ from each other or from control plants. In contrast, plants inoculated with 20,000 RKN eggs and all multi-pathogen infections had significantly reduced pod yields either in terms of number or weight. Effect of combined inoculation of nematode and virus on seed yield in experimental field and on-farm trials. At Ibadan, Malayan soybean plants inoculated with the virus alone produced significantly lower seed yields than plants inoculated with RKN (Table 1). When nematode inoculation was followed by virus inoculation one week later, the combined infection resulted in lower yields than single infections with either the virus or nematode. However, nematode inoculation followed by virus inoculation 2 weeks later did not significantly reduce yield relative to plants inoculated with SMV alone, although it reduced yield compared to plants inoculated with RKN alone. In TGX 923-2E, seed yields in virus-inoculated plants did not differ significantly from those inoculated with nematode followed by the virus one week later, but yields were significantly lower in treatments inoculated first with nematode followed by the virus 2 weeks later. Combined inoculations by both pathogens reduced seed yield more than single inoculation with the nematode. In TGM 80, combined inoculations significantly reduced seed yield more than single inoculations with either virus or nematode. Likewise, combined inoculation reduced yield more than single inoculations in TGM 1784. At Ayepe, the trend was similar to that at Ibadan (Table 1). With the exception of plants inoculated with the nematode alone, all other treatments resulted in significantly lower seed yields compared to the uninoculated control. Single inoculations by either the virus or nematode did not reduce yield as much as combined inoculations, irrespective of cultivar. DISCUSSION The greenhouse study clearly showed that, in general, simultaneous inoculation with SMV and RKN significantly reduced the numbers, and fresh and dry weights of pods as compared with uninoculated control plants or plants inoculated only with RKN (Fig. 1). This was irrespective of the reaction of individual varieties to the virus or nematode (Iheukwumere et al., 1995). It was also apparent that in soybean cv. TGM 1784, which is resistant to both virus and nematode individually (Iheukwumere et al., 1995), inoculation of SMV in combination with any RKN inoculum level resulted in reduced pod yields. On the other hand, for varieties less resistant to either the virus or nematode, multi-pathogen infection resulted in yields lower than those from nematode-inoculated plants only when the nematode inoculum level reached 20,000 eggs per plant. This was an indication that multi-pathogen infection by SMV and RKN predisposes resistant plants to more severe damage from the nematodes. This was further supported in combined or successive inoculation of the nematode and virus in the field trials (Table 1). Increased pathogenicity of M. incognita when in multi-pathogen infection with tobacco ring spot virus has also been demonstrated for soybean (Rhyder and Crittenden, 1962). This effect has similarly been reported in plants species as varied as maize and tobacco (Nicotiana tobacun L.) when they are infected by species of Meloidogyne and a mosaic virus (Bird, 1969; Khurana et al., 1970). In the greenhouse study, and to some extent in the field trial, SMV inoculation alone caused as much damage as inoculation with the virus and RKN in most yield and yield components. This was, however, not the case in plants inoculated with RKN alone, although there was an obvious increase in severity as the inoculum level increased to 20,000 eggs per plant. It is probable that some nematode inoculation levels beyond this point may produce the same effect as in multi-pathogen or single infection with virus, but this was not investigated. This is because the severity of damage in RKN-infected plants is often related to the initial population density or number of juveniles penetrating and becoming established within the root tissues of young plants (Schmitt and Noel, 1984; Hussey, 1985). In the field studies, multi-pathogen inoculation caused more severe damage as indicated by a significant reduction in seed yield, than nematode inoculation alone and the uninoculated control treatments. However, in terms of yield, multi-pathogen infections were generally not different from single infection with the virus (Alam et al., 1990). These differences may be due to the conditions under which the studies were performed. Whereas most reports were based on the results of greenhouse or screen house tests, our studies included field trials. Although field trials are more likely to represent the situation in actual farmerselds, environmental conditions are more variable, and thus more difficult to control. For instance, colonising and transient aphid vectors of SMV could introduce the virus from sources outside the field (Halbert et al., 1981). Since mixed infections by the virus and nematode generally caused a more severe yield reduction than single infections of the virus under all conditions in this study, future research should be directed towards evaluating or breeding for combined resistance to both pathogens in soybean. ACKNOWLEDGEMENTS The authors wish to thank the International Institute of Tropical Agriculture and International Foundation for Science for financial support. REFERENCES Adesiyan, S.O., Caveness, F.E., Adeniji, M.O. and Fawole, B. 1990. Nematode Pests of Tropical Crops. Heinamann, Ibadan. 144 pp. Alam, M.M., Samad, A. and Anver, S. 1990. Interaction between tomato mosaic virus and Meloidogyne incognita. Nematologia Mediterranea 18:131-133. Anon. 1990. Soybeans for Good Health. International Institute of Tropical Agriculture, Ibadan. 23 pp. Bird, A.F. 1969. The influence of tobacco mosaic virus on the growth of Meloidogyne javanica. Nematologica 15:201-209. Chen, P., Buss, G.R., Roane, C.W. and Tolin, S.A. 1994. Inheritance in soybean of resistant and necrotic reactions to soybean mosaic virus strains. Crop Science 34:414-422. Eisenback, J.D., Hirschmann, H., Sasser, J.N. and Triantaphyllou, A.C. 1981. A Guide to the Four Most Common Species of Root-Knot Nematodes (Meloidogyne Spp.) with a Pictorial Key. North Carolina State University Graphics, Raleigh. 48 pp. Fehr, W.R. and Caviness, C.E. 1977. Stages of Soybean Development. Iowa Agriculture and Home Economics Experiment Station Special Report 80. Gomez, K.A. and Gomez, A.A. 1994. Statistical Procedures for Agricultural Research. John Wiley, New York. 680 pp. Goswami, B.K. and Chenulu, V.V. 1974. Interaction of root-knot nematode, Meloidogyne incognita, and tobacco mosaic virus in tomato. Indian Journal of Nematology 4:69-80. Halbert, S.E., Irwin, M.E. and Goodman, R.M. 1981. Alate aphid (Homoptera: Aphididae) species and their relative importance as field vectors of soybean mosaic virus. Annals of Applied Biology 97:1-19. Hartman, K.M. and Sasser, J.N. 1985. Identification of Meloidogyne species on the basis of differential host test and perennial-pattern morphology. In: An advanced Treatise on Meloidogyne, Volune 11: Methodology. Barket, K.R., Carter, C.C. and Sasser, J.N. (Eds.), pp. 69-78. North Carolina State University Graphics, Raleigh. Hussey, R.S. and Barker, K.R. 1973. A comparison of methods of collecting of Meloidogyne spp., including a new technique. Plant Disease Reporter 57:1025-1028. Iheukwumere, C.C., Atiri, G.I., Fawole, B. and Dashiell, K. 1995. Evaluation of some commonly grown soybean cultivars for resistance to the root-knot nematode and soybean mosaic virus in Nigeria. Fitopatologia Brasileira 20:190-193. Khurana, S.M.P., Goswami, B.K. and Raychaudri, S.P. 1970. Interaction of maize mosaic with root-knot nematode Meloidogyne incognita (Kofoid and White) Chitwood in maize (Zea mays L.). Phytopathologiche Zeitschrift 69:267-272. Mutsaers, H.J.W. 1991. Opportunities for Second Season Cropping in South-western Nigeria. Research and Crop Management Programme monograph No. 4, International Institute of Tropical Agriculture, Ibadan, Nigeria. 28 pp. Rhyder, H.W. and Crittenden, H.W. 1962. Interrelationship of tobacco ring spot virus and Meloidogyne incognita acrita, in roots of soybeans. Phytopathology 52:165-166. Riggs, R.D. and Schmitt, D.P. 1987. Nematodes. In: Soybeans Improvement, Production and Uses. Wilcox, J.R. (Ed.), pp. 757-778. American Society of Agronomy, Madison. Rossel, H.W. and Thottappilly, G. 1985. Virus Diseases of Important Food Crops in Tropical Africa. International Institute of Tropical Agriculture, Ibadan. 61pp. Schmitt, D.P. and Noel, G.R. 1984. Nematode parasites of soybeans. In: Plant and Insect Nematodes. Nickel, W.R. (Ed.), pp. 14-59. Marcel Dekker, New York. Thottappilly, G. 1992. Plant virus diseases of importance to African Agriculture. Journal of Phytopathology 134:265-268. Walkey, D.G.A. 1991. Applied Plant Virology. Chapman and Hall, London. 338 pp. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Photo images[cs96075b.jpg]Line drawing images[cs96075a.gif] |
| |||||||||

{kind=link}
{kind=link}