 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Short communication : phosphorus, sulphur and molybdenum effects on response of Phaseolus beans to inoculation with Rhizobium in Malawi
N. Z. LUPWAYI1 and A. B. C. MKANDAWIRE Crop Science Department, University of Malawi, Bunda College of Agriculture, P. O. Box 219, Lilongwe, Malawi (Received 21 September, 1995; accepted 19 September, 1996)
Code Number: CS96076
Sizes of Files:
Text: 19.7K
Graphics: Tables (jpg) - 250K ABSTRACT The effects of P, S and Mo on the response of field beans (Phaseolus vulgaris L.) to inoculation with Rhizobium leguminosarum bv. phaseoli were investigated at four sites in Malawi. Treatments consisted of non-inoculation and inoculation; fertilisation at 0 and 17.5 kg P ha^-1; 0 and 30 kg S ha^-1; and 0 and 1 kg Mo ha^-1. A nitrogen control at 100 kg N ha^-1 was included. To identify the nutrients that limited response to inoculation most, P, S and Mo were either applied or not applied to the inoculated plants. At all study sites, there was no significant response (P < 0.05) of nodule number to inoculation. Grain yield increased by 39% due to inoculation at Champhira. Improved soil fertility increased nodule number by 387 to 426% at three of the four sites. Also, significant increases (31% and 35%) in grain yield due to improved soil fertility occurred at two sites. At least one of the three nutrients was identified as limiting nodulation at each site, but grain yields were only affected by P deficiency at Sokola and Mo deficiency at Champhira. Therefore, alleviating nutrient deficiencies and increasing nodule occupancies by efficient inoculant strains are some of the ways in which the response of beans to inoculation could be increased. Key Words: Field beans, nitrogen fixation, nodulation, soil fertility
RESUME
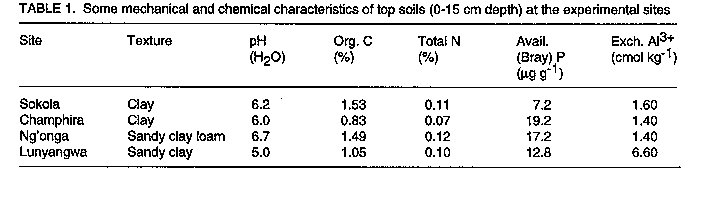
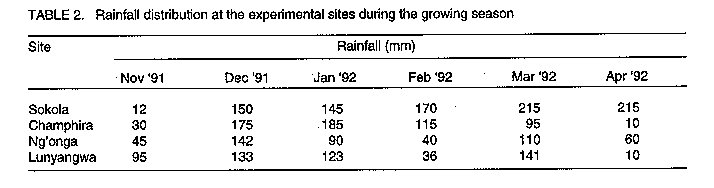
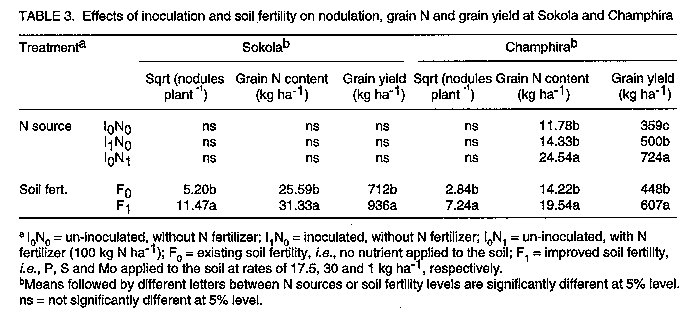
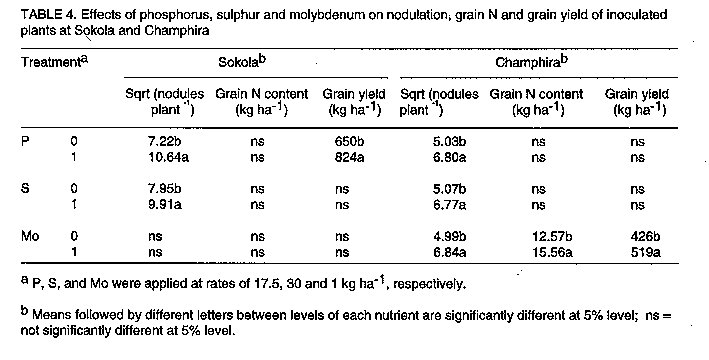
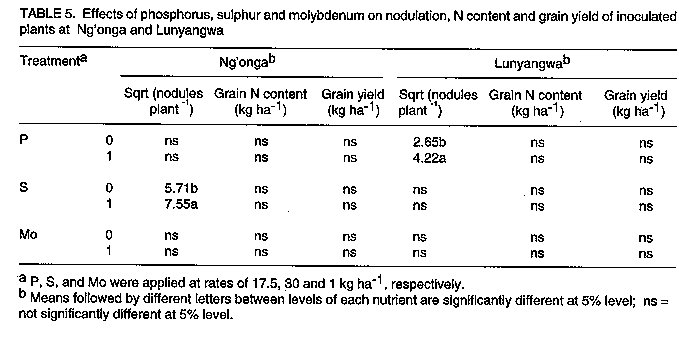
Les effets de la reaction des P, S et Mo dans les champs d'haricot (Phaseolus Vulgaris L.) en inoculation avec le Rhizobium Leguminosarum par phaseoli etaient etudies dans six centres au Malawi. Les traitements qui ont consiste a la non-inoculation et a l'inoculation; la fertilisation entre 0 et 17.5 kg P par hectare^-1; 0 et 30 kg S par hectare^-1, et 1 kg Mo l'hectare. Un systeme de controle du nitrogene a 100 kg N par hectare-1 etait inclus. Pour identifier les elements nutritifs qui ont limite la reaction a l'inoculation, P, S et Mo etaient soit appliques ou non appliques aux plantes inoculees. Dans tous les centres d'etudes, il n'y avait pas de reactions sensibles (P<0.05) du nombre de nodule a l'inoculation. Lerendement de graines a augmente de 39% grece a l'inoculation a Champira. Le changement de la fertilite du sol a augmente le nombre de nodules de 387 a 426% dans trois centres sur quatre. Il y a aussi eu, d'augmentations importantes (entre 31% et 35%) en productions des grains a cause du changement de la fertilite du sol qui a eu lieu dans deux sites. Plus au moins l'un des trois elements nutritifs etait identifie comme une nodulation qui se limite a chaque site, mais les rendements des grains etait identifie comme une nodulation qui se limite a chaque site, mais les rendemets des grains etaient seulement affectes par la deficience P a Sokola et la deficience M a Champira. Par consequent, pour alleger les elements nutritifs des deficiences et augmenter les occupations des nodules par les lignees effectives des inoculants sont les moyens par lesquels les reaction dans les champs d'haricot a l'inoculation pourraient etre augmentes. Mots Cles: Champ d'haricot, fixation du nitrogene, nodulation, fertilite du sol INTRODUCTION Inoculation of field beans (Phaseolus vulgaris L.) with rhizobia rarely increases grain yields (Graham, 1981; Davis, 1982). For this reason, application of N fertilizer is usually recommended for bean production; for example, 46 kg N ha^-1 in Malawi (Ministry of Agriculture, 1990). However, because most smallholder farmers cannot afford N fertilizer, beans are usually N deficient and produce poor yields. Exploiting biological nitrogen fixation (BNF) through inoculation with rhizobia would be a cheaper option than applying N fertilizer, especially if high N-fixing combinations of rhizobia and bean genotypes could be identified. Beans are known to fix little amounts of N (LaRue and Patterson, 1981). However, inoculation with some strains of rhizobia has produced bean yields as high as those obtained when 180 kg N ha^-1 was applied (2.2 t ha^-1 inoculated and 2.4 t ha^-1 N-fertilized) (Kipe-Nolt and Pineda, 1988). Several factors including rhizobium, plant genotype and environment limit N fixation (Keyser and Li, 1992). One important environmental factor that affects both the rhizobium and the host plant is the nutrient status of the soil. Phosphorus is known to be among the most limiting factors of BNF in tropical soils in Africa (Keya, 1977; Haque et al., 1996), Latin America (Hernandez et al., 1982) and Australia (Andrew and Jones, 1978). Sulphur (Edwards, 1977) and molybdenum (Robson, 1978; de Yunda and de Gonzalez, 1982) are also important nutrients in BNF. They are both components of nitrogenase, the N-fixing enzyme of rhizobia (Pau, 1991). It is not clear to what extent deficiencies of these nutrients limit the response of field beans to rhizobia inoculation in Malawi soils. Therefore, experiments were conducted to investigate the effects of P, S and Mo on the response of Phaseolus beans to inoculation. MATERIALS AND METHODS A field experiment was conducted at Sokola (09 degrees 40' S, 33 degrees 20' E) in Chitipa District; Ng'onga (10 degrees 40' S, 33 degrees 40' E) in Rumphi District; Lunyangwa (11 degrees 30' S, 34 degrees 10' E) in Mzimba District; and Champhira (12 degrees 20' S, 33 degrees 30' E) in Mzimba District. Soil properties at these sites are presented in Table 1 and rainfall data for the season are shown in Table 2. All the sites had been under fallow for at least two years. At each site, the experiment consisted of two parts on the same plot, each replicated three times. One part, which was aimed at evaluating response of beans to inoculation and soil fertility, consisted of two factors arranged in a split-plot design. Treatments consisted of N source as a main-plot factor:- non-inoculated, without fertilizer N (I0N0); inoculated, without fertilizer N (I[1]N[0]) and non-inoculated, with fertilizer N at 100 kg N ha^-1 (I[0]N[1]). Nitrogen was applied as urea in split applications; one third of the total rate was applied at planting and the remaining two thirds 21 days after planting. Soil fertility was the sub-plot factor and included:- existing soil fertility, i.e., no nutrient applied to the soil (F[0]); and improved soil fertility, i.e., P, S and Mo applied to the soil (F[1]). The second part of the experiment was designed to investigate which of the applied nutrients, P, S or Mo, affected the performance of inoculated plants most. Thus, inoculated plants (I[1]N[0]) were grown with or without applied P, S and Mo. A randomised complete block design was used. In their respective treatments, P at 17.5 kg P ha^-1 was applied as triple superphosphate, S at 30 kg S ha^-1 as gypsum, and Mo at 1 kg Mo ha^-1 as ammonium molybdate, all at planting time. Seeds of P. vulgaris genotype CIAT25-2 (a Type 2 genotype, i.e., intermediate between dwarf and climbing types), were planted 0.1 m apart in single rows on ridges 0.9 m apart and 4 m long. The crops were planted in December, 1991. Inoculation was done with peat-based inoculant of Rhizobium leguminosarum bv. phaseoli strain MG336, recommended for field beans in Malawi. The seeds were inoculated at a rate of 1 x 10^5 cells per seed immediately before planting. At 50 percent flowering, nodules were counted on eight plants per treatment. Grain harvesting was done at physiological maturity and yield was recorded at 12% moisture content. Grain N content was analysed using the Kjeldahl technique (Anderson and Ingram, 1993). Since about 80% of N in grain legumes is found in the grain at harvest (Eaglesham et al., 1982; Chandel et al., 1989; Jefing et al., 1992), grain N content is a good indicator of total plant N content. Analysis of variance suitable for the experimental design of each experimental part was performed on the data, using MSTAT-C computer software (Michigan State University, 1988). Nodule number data were transformed to square roots in all the analyses in order to homogenise their variances. RESULTS At Sokola, neither inoculation (I[1]N[0]) nor N fertilizer (I[0]N[1]) significantly influenced nodulation, grain N or grain yield (P > 0.05)(Table 3). Improved soil fertility (F[1]), however, increased nodulation by 387% (using untransformed data), grain N by 22% and grain yield by 31% (Table 3). Table 4 shows that P and S were the limiting nutrients for nodulation; in addition, P limited grain yield. There were no significant interactions between N source and soil fertility level, or among the three nutrients. At Champhira, nodule number and grain N were not affected significantly by inoculation, but grain yield increased by 39% with inoculation and 102% with N fertilizer (Table 3). With improved soil fertility, nodulation, grain N and grain yield increased by 547%, 37% and 35%, respectively. Table 4 shows that P, S, and Mo all limited nodulation, and Mo limited grain N and grain yield too. No significant interactions between N source and fertility level, or among the three nutrients, were detected. At Ng'onga and Lunyangwa, inoculation and N fertilizer had no significant effects on nodule number, grain N or grain yield (data not shown). Improved soil fertility had no significant effect on the measured parameters at Ng'onga, although Table 5 shows that S limited nodulation. Surprisingly, S increased nodulation more without than with Mo application (data not shown). At Lunyangwa, improved soil fertility increased nodule number by 426% (from 3.4 to 17.9 nodules plant-1), mainly due to application of P. No interactions were detected at this site either. DISCUSSION Nodulation, grain N content and grain yield were very low at Lunyangwa. A combination of factors including soil acidity (high exchangeable Al^3+, Table 1), drought during the season (Table 2) and infestation by bean fly (Ophiomyia spp.) (field observations) all contributed to the poor performance. Liming is recommended at this site. There was no response of nodule number to inoculation at all sites (Table 3). This is a reflection of a large population of indigenous soil rhizobia capable of nodulating beans. Since application of N fertilizer increased grain N and grain yield relative to non-inoculation in some cases, it can be concluded that indigenous rhizobia are inefficient fixers of nitrogen. In such situations, competition for nodule formation between inoculant strains and indigenous rhizobia becomes a very important factor in the response of a legume to inoculation (Streeter, 1994). Analysis of nodule occupancy is necessary to find out the proportion of nodules formed by inoculant strains. Several factors are known to affect nodule occupancy (Streeter, 1994). Some of them, such as dose, formulation and placement of inoculant, can be manipulated easily to increase nodule occupancy. Nodulation increased markedly with improved soil fertility (Table 3). At least one of the three nutrients limited nodulation at each site and, whenever more than one nutrient was limiting, one of them was P (Tables 4 and 5). This indicates that the natural fertility of these soils is a limiting factor to the nodulation process. However, it was only at Sokola and Champhira that the increase in nodulation with improved soil fertility was associated with increased grain yield. Table 2 shows that the rainfall distribution at these two sites was more even during the season than at Ng'onga and Lunyangwa which received little rain in the middle of the season. Dry soil conditions may partly explain why grain yield did not respond to improved soil fertility at Ng'onga and Lunyangwa. The other problems at Lunyangwa mentioned above equally applied in this case. Grain yield response to application of P fertilizer at Sokola is in agreement with soil analysis results which indicate low P content (Table 1). At Champhira, Mo clearly limited bean productivity as application of this nutrient significantly increased nodulation, grain N and grain yield. Low soil fertility as a constraint to bean production has been identified in other bean-growing areas in Africa. In Uganda, multi-location yield-loss-assessment trials were conducted in farmers' fields for two seasons to assess the effects of fertilisation with N and P, and control of beanfly and some fungal diseases. The greatest yield reduction at all sites was due to N and P deficiency (CIAT, 1990). In similar trials in Tanzania, low soil fertility was a major constraint at one (Lushoto) of three sites (CIAT, 1990). Phosphorus deficiency is a critical constraint to production of beans and other crops in tropical regions, yet most small-scale farmers cannot afford P fertilizers. One approach to alleviate this problem is to identify bean genotypes that are tolerant to low soil P levels. Use of rock phosphates, which are usually cheaper than refined superphosphates, is another alternative in raising the P status of soils. Evaluation of the agronomic and economic effectiveness of rock phosphates for bean production is recommended. Farmyard manure, crop residues and leaf prunings, should be applied whenever possible since they supply multiple nutrients to the soil concurrently. In conclusion, nodulation by indigenous rhizobia and low soil fertility, especially P deficiency, have been found to be limiting factors to the effectiveness of inoculants in bean production. Therefore, alleviating nutrient deficiencies and pH, and increasing nodule occupancies by efficient inoculant strains are some of the ways in which the response of beans to inoculation could be improved.
ACKNOWLEDGMENTS
Field technical assistance by Mr D.S. Jere of the Malawi National Bean Programme is highly appreciated. This study was partly funded by the USAID Bean/Cowpea Collaborative Research Support Programme. REFERENCES Anderson, J.M. and Ingram, J.S.I. (Eds.). 1993. Tropical Soil Biology and Fertility: A Handbook of Methods. 2nd Edition. C.A.B. International, Wallingford. 221pp. Andrew, C.S. and Jones, R.K. 1978. The phosphorus nutrition of tropical forage legumes. In: Mineral Nutrition of Legumes in Tropical and Subtropical Soils. Andrew, C.S. and Kamprath, E.J. (Eds.), pp. 295-311. CSIRO, Melbourne. Chandel, A.S., Pandey, K.N. and Saxena, A.C. 1989. Symbiotic nitrogen fixation and nitrogen benefits by nodulated soybean (Glycine max (L.) Merrill) to interplanted crops in northern India. Tropical Agriculture (Trinidad) 66:73-77. CIAT (Centro Internacional de Agricultura Tropical). 1990. Bean Programme Annual Report, 1989. CIAT. Cali, Colombia. Davis, P.E. 1982. Legume Microbiology Research in Malawi, 1976 to 1981. Final Report for the ODA Technical Cooperation Officer. Ministry of Agriculture, Lilongwe, Malawi. Eaglesham, A.R.J., Ayanaba, A., Ranga Rao, V. and Eskew, D.L. 1992. Mineral N effects on cowpea and soybean crops in a Nigerian soil. II. Amounts of N fixed and accrual to the soil. Plant and Soil 68:183-192. Edwards, D.G. 1977. Nutritional factors limiting nitrogen fixation by rhizobia. In: Biological Nitrogen Fixation in Farming Systems of the Tropics. Ayanaba, A. and Dart, P.J. (Eds.), pp. 189-204. John Wiley and Sons, Chichester. Graham, P.H. 1981. Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris L.: a review. Field Crops Research 4:93-112. Haque, I., Lupwayi, N.Z. and Luyindula, N. 1996. Inoculation and phosphorus effects on Desmodium intortum and Sesbania sesban in the Ethiopian highlands. Agriculture, Ecosystems & Environment 56:165-172. Hernandez, B.C., Mendez-Lay, J.M. and Focht, D.D. 1982. Nitrogen and phosphorus requirements for the growth and nodulation of Cajanus cajan in Panamanian soils. In: Biological Nitrogen Technology for Tropical Agriculture. Graham, P.H. and Harris, S.C. (Eds.), pp. 167-172. Centro Internacional Agricultura de Tropical (CIAT), Cali, Colombia. Jefing, Y., Herridge, D.F., Peoples, M.B. and Berkasem, B. 1992. Effects of N fertilization on N2 fixation and N balances of soya bean grown after lowland rice. Plant and Soil 147:235-242. Keya, S.O. 1977. Nodulation and nitrogen fixation in legumes in East Africa. In: Biological Nitrogen Fixation in Farming Systems of the Tropics. Ayanaba, A. and Dart, P.J. (Eds.), pp. 233-244. John Wiley and Sons, Chichester. Keyser, H.H. and Li, F. 1992. Potential for increasing biological nitrogen fixation in soybean. Plant and Soil 141:119-135. Kipe-Nolt, J. and Pineda, P. 1988. A strategy to improve nitrogen fixation: Examples from El Salvador. Bean Newsletter 10:8-10. LaRue, T.A. and Patterson, T.G. 1981. How much nitrogen do legumes fix? Advances in Agronomy 34:15-37. Michigan State University. 1991. MSTAT-C, A Microcomputer Programme for the Design, Management and Analysis of Agronomic Research Experiments. Michigan State University, Michigan, USA. Ministry of Agriculture. 1990. Guide to Agricultural Production in Malawi, 1990-1991. Ministry of Agriculture, Lilongwe, Malawi. Pau, R.N. 1991. The alternative nitrogenases. In: Biology and Biochemistry of Nitrogen Fixation. Dilworth, M.J. and Glenn, A.R. (Eds.), pp. 37-57. Elsevier, Amsterdam. Robson, A.D. 1978. Mineral nutrients limiting nitrogen fixation in legumes. In: Mineral Nutrition of Legumes in Tropical and Subtropical Soils. Andrew, C.S. and Kamprath, E.J. (Eds.), pp. 277-293. CSIRO, Melbourne. Streeter, J.G. 1994. Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Canadian Journal of Microbiology 40:513-522. de Yunda, A.L. and Gonzalez, N.M. 1982. Influence of molybdenum on nitrogen fixation by white clover in the Bogota savanna. In: Biological Nitrogen Technology for Tropical Agriculture. Graham, P.H. and Harris, S.C. (Eds.), pp. 161-166. Centro Internacional Agricultura de Tropical (CIAT), Cali, Colombia. Copyright 1996 The African Crop Science Society
The following images related to this document are available:Photo images[cs96076b.jpg] [cs96076d.jpg] [cs96076a.jpg] [cs96076e.jpg] [cs96076c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}