 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Estimates of genetic variances and interrelationships of traits associated with Striga resistance in maize L. Akanvou, E. V. Doku^1 and J. Kling^2 IDESSA, B.P. 121 Ferkessedougou, Cote d'Ivoire
^1 University of Ghana, Crop Science Department, P.O. Box 44 Legon, Accra,
Ghana (Received 19 September, 1996; accepted 31 December, 1996)
Code Number: CS97001
Sizes of Files:
Text: 24.5K
Graphics: Tables (gif) - 29.3K
ABSTRACTThe choice of the selection method to be used in the genetic improvement of a crop depends upon the type of gene action controlling the characters under study. Design 1 crosses were made in an open-pollinated maize population, TZL Composite-1 C1. As a result, 320 full-sib families were generated and tested under artificial infestation of Striga at two locations, Mokwa and Abuja in Nigeria. Study of gene action involved in the inheritance of S. hermonthica resistance was done. Additive genetic variance was a major portion of the genotypic variance for characters like ear Striga rating (sigma 2a = 0.41+/-0.15 and sigma 2d = -0.20+/- 0.23), yield of infested plants (sigma 2a = 401564+/-172649 and sigma 2d = 57127+/-315131) and for the Striga syndrome rating (sigma 2a = 0.28+/-0.12 and sigma 2d = 0.03+/-0.21). However, the number of Striga plants supported by the host was controlled by non-additive gene action (sigma 2a = 75.69+/-76.64 and sigma 2d = 211.47+/-134.48). Grain yield was positively correlated to the number of ears harvested (rg = 0.99+/-1.12), but negatively correlated to Striga syndrome rating at 8 weeks after planting (rg = -0.92+/-0.93), ear Striga rating at harvest (rg = -0.88+/-1.28), and to a lesser extent, to Striga counts at 8 weeks (rg = -0.22+/-0.46) and at 10 weeks (rg = -0.09+/-0.75) after planting. Key Words: Gene action, genetic correlation, Striga hermonthica RESUME Le choix de la methode de selection a adopter pour une amelioration varietale depend du mode d'action de genes qui controlent les caracteres etudies. Des croisements de type Design I ont ete faits dans une population de mais a pollinisation libre, TZL Composite-1 C1. Ainsi, 320 familles full-sibs ont ete generes et testes sous infestation artificielle avec un parasite de mauvaises herbes, Striga [Striga hermonthica (Del.) Benth.] a deux localites, Mokwa et Abuja au Nigeria. L'etude de l'action des genes impliques dans la transmission de la resistance au S. hermonthica ete faite. La variance additive representait une importante proportion de la variance genetique pour les caracteres tels que le syndrome du Striga sur les epis (sigma 2a = 0.41+/-0.15 et sigma 2d = -0.20+/-0.23), le rendement des plantes infestees (sigma 2a = 401564+/-172649 et sigma 2d = 57127+/-315131) et pour le syndrome du Striga visible sur la plante (sigma 2a = 0.28+/-0.12 et sigma 2d = 0.03+/-0.21). Cependant, le nombre de plants de Striga supporte par la plante hote est controle par une action de genes non-additive (sigma 2a = 75.69+/-76.64 et sigma 2d = 211.47+/-134.48). Les correlations genetiques entre les caracteres associes avec la resistance au Striga ont ete estimes. Le rendement etait positivement correle au nombre d'epis recoltes (rg = 0.99+/- 1.12), mais negativement correle au syndrome du Striga 8 semaines apres semis (rg = -0.92+/-0.93), au syndrome du Striga sur l'epis a la recolte (rg = -0.88+/-1.28), et a un degre moindre au nombre de plantes de Striga 8 semaines (rg = -0.22+/-0.46) et 10 semaines (rg = -0.09+/- 0.75) apres semis. Mots Cles: Action des genes, correlation genetique, Striga hermonthica INTRODUCTION Maize (Zea mays L.) is one of the most widely grown cereals in Africa. It is utilised for human consumption, animal feed and as industrial raw material. Despite the increasing demand for maize, its production per unit area is not sufficient to meet the demands of a rapidly growing African population. Diseases, insects, and parasitic weeds especially Striga [Striga hermonthica (Del.) Benth.] are the major causes of low yields. Striga, the most widely distributed species, is a root parasitic plant of the Scrophulariaceae family and is known as one of the major constraints to maize production in sub-Saharan Africa. In maize, crop losses due to Striga vary between 10% to total crop yield failure (Smaling et al., 1991). Control methods using herbicides or nitrogen fertilizers are costly for most African farmers. On the other hand, hoeing or hand-pulling of Striga is time consuming, very tedious and not effective. Breeding for resistant varieties, combined with legumes/crop rotation, are among the most effective and economical approaches for solving the Striga problem. Therefore, knowledge of the type of gene action involved in Striga resistance will help in defining effective breeding strategies. The objectives of this research were to: (i) investigate the type of gene action involved in the inheritance of Striga resistance in the open-pollinated maize population, TZL Composite 1-C1, and (ii) estimate genetic correlations among traits associated with Striga resistance. MATERIAL AND METHODS Experimental design. A bulk of the maize population TZLComp1-C1 was planted at the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria and North Carolina Design 1 crosses were made: 80 random plants designated as male were each crossed with 4 other random plants designated as female. Seeds for plants used as males were planted 4 days after planting the female parents. Every plant used as a male parent was selfed. At harvest, a set of 4 female ears representing a cross to a particular male were selected and constituted a single male group (half-sib family). A total of 80 such male groups were obtained, resulting in 320 full-sib families which were tested under Striga artificial infestation in a randomised complete block design with two replications at two locations, Abuja and Mokwa (Northern Nigeria). The 320 full-sib families were divided into 20 sets which contained 16 families and also 2 hybrid checks (9022-13 and 8338-1) each. A total of 18 entries were assigned to each plot. Each plot consisted of two rows of 22 plants. One row of each plot was artificially infested with Striga while the other row remained non-infested. The rows were 5 m long and 0.75 m apart, with plants spaced 50 cm within rows. Three kernels per hill were planted and the stand thinned to 2 plants per hill. Infested and non-infested plots were separated by 2m alleys to avoid contamination. Striga infestation and field management. Screening for Striga resistance in population TZLComp1-C1 was done on artificially infested fields at Abuja and Mokwa. In both locations, maize was planted on ridges which provided enough moisture and prevented waterlogging. Fertilizer NPK (15:15:15) was applied to each row after planting at a rate of 60 kg ha^-1. The Nitrogen rate, which was half of the recommended rate for maize in Northern Nigeria soils, was used to ensure maximum development of Striga. Half of the N fertilizer (30 kg ha^-1) was applied at planting and the other half was top-dressed 4 to 5 weeks later. The experiment was kept free of weeds other than Striga.
Striga seeds were mixed with sieved sand which was used as a carrier material to provide adequate volume for rapid and consistent infestation. The amount of sand and Striga seeds needed for artificial infestation was calculated following the method outlined by Berner et al. (1993). Prior to Striga infestation and maize planting, ethylene gas was used to stimulate suicidal germination of existing Striga seeds in the soil. A scoop that held approximately 5 ml of water was used for infestation. Holes of 10 cm in diameter and 8 cm in depth were dug out on the ridges. Infestation was then done by pouring the content of a scoop filled with Striga seed mixed with sieved sand in each of the holes of the infested rows. About 3,000 germinable Striga seeds were sowed in each hole. Striga seeds were then lightly covered with surrounding soil. Maize seeds were planted the same day, both in the non -infested hills and infested hills above the Striga seeds. Data collection. Data were collected on infested and non-infested rows for the following parameters: date of flowering, plant height, ear height, number of Striga plants emerged per row at 8 weeks after planting (Striga count 1), Striga syndrome on host plants at 8 weeks after planting (Striga rating 1), number of ears per row, Striga effect on ears (ear Striga rating) and yield was estimated. Date of flowering was recorded when approximately 50% of the plants were shedding pollen. Plant and ear height were measured from ground level to the tip of the tassel and uppermost ear bearing node, respectively. Data on the number of Striga plants emerged and Striga syndrome on host plants were taken on the entire infested rows. Plant aspect was taken on host plants in non-infested rows only. The number of ears was recorded for each infested and non-infested row. Moisture was measured on infested and non-infested rows by using a moisture meter (Dicky-John Corp., U.S.A). Estimates of moisture were determined on samples of grains shelled from the middle areas of at least 5 ears for each row. Yield of infested and non-infested rows was computed in kg ha^-1. The Striga syndrome rating (1 = no symptoms, 9 = severe symptoms) used to measure tolerance of maize to Striga was based on several indicators: Foliar damage which increases with the severity of Striga infestation (Small 1-3 mm, round, white chlorotic blotches, gradually extending longitudinally into streaks); loss of turgor; and in severly affected plants Ôscorching' or firing of the leaf tissue, particularly around the margins. Affected plants exhibited spindly and weak stems, small and poorly filled ears, shortening of internodes which gave a dwarfing appearance. This scale was first used at IITA by Kim and Winslow (1991). Statistical analysis. Data for each plot were used for analysis of variance (ANOVA) for each character combined for the two locations using Proc GLM from SAS (SAS, 1987). Pooled analysis over sets for both locations was done according to the method of Hallauer and Miranda (1988). Estimate of the components of variance and their standard errors. By using the appropriate mean squares obtained from the analysis of variance (Table 1), an estimate of the variance among males (sigma 2m), and variance among females mated to the same male (sigma 2f), were computed according to the formula of Lindey et al. (1962).
Under the assumptions outlined by Comstock and Robinson (1948), additive variance (sigma 2a) and non-additive genetic variance, denoted as dominance variance (sigma 2d) were estimated as follows:
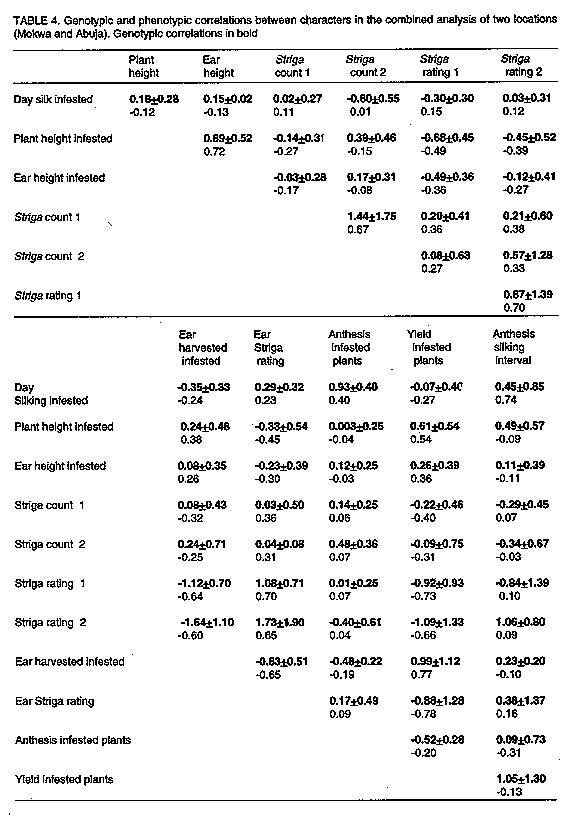
Estimate of genotypic and phenotypic correlations. Genotypic correlations (rg) and phenotypic correlations (rp) were calculated using the mean products and estimates of genetic and phenotypic variances as outlined by Falconer (1981). The standard error (S.E) of the genetic correlations were computed using the formula given by Scheinberg (1966). RESULTS Estimate of the components of variance and their standard errors. From the analysis of variance (ANOVA), sigma 2m and sigma 2f were computed (Table 2). Comparison of the estimates of variances sigma 2a and sigma 2d (Table 3) showed that the estimates of sigma 2a were lower than those of sigma 2d for Striga count 1, yield uninfested and delay silking. Estimates of sigma 2a were greater than that of sigma 2d for Striga rating1, ear Striga rating, yield of infested plants and for anthesis silk interval of infested plants. In the case of the yield of non-infested plants, sigma 2a and sigma 2d were similar in magnitude. Table 3 also showed that the ratio sigma 2d/sigma 2a was less than 1.0 for Striga rating 1 (0.135) and yield of infested plant (0.142) and was slightly greater than 1.0 for yield of un-infested plants (1.114). On the other hand, the ratio was considerably greater than 1.0 for Striga count 1 (2.79) and for delay silking assuming sigma 2a was a value near zero.
Estimates of phenotypic and genotypic correlations. Correlations and their standard errors are presented in Table 4. Positive but low genotypic correlations were obtained between Striga count 1 and Striga rating1 (rg = 0.20+/-0.41), Striga count 1 and ear Striga rating (rg = 0.036+/-0.50), Striga count1 and day silking for infested plants (rg = 0.020+/-0.27) and Striga count1 and anthesis (rg = 0.14+/-0.25). Positive and high genotypic correlations were obtained between Striga rating 1 and ear Striga rating (rg = 1.08+/-0.71), Striga rating 1 and Striga rating 2 (rg = 0.87+/-1.39) and anthesis of infested plants and day silking of infested plants (rg = 0.93+/-0.40). These high values of genotypic correlations indicated that there may be considerable genetic association between the tested characters. Yield showed a high genotypic correlation with plant height (rg = 0.61+/-0.54) and with ears harvested for infested plants (rg = 0.99+/- 1.12), which is a component of yield. Negative correlations were obtained between Striga count 1 and yield (rg = -0.227 +/- 0.46), Striga rating and yield (rg = -0.92 +/- 0.93) and ear Striga rating and yield (rg = -0.88 +/- 1.28). Such negative associations were expected since Striga affects the physiology and the yield of infested plants. DISCUSSION Comparative magnitude of the additive genetic variance and dominance variance. A two factor design (Design 1) which generated full-sib families within half-sib families was used in this study to detect the presence of genetic variability. Design 1 only permits the estimation of genetic components of variance such as additive genetic variance and dominance variance which might also contain epistatic variances if there is epistasis. However, several variance estimation studies (Compton et al., 1965; Eberhart et al., 1966; Stuber and Moll, 1969, 1971) suggested that epistatic variability is negligeable in maize, both within open-pollinated varieties and in varietal hybrids (F1 crosses of open-pollinated varieties). Negative genetic variance component estimates obtained in Table 3 could have been due to sampling error or lack of random mating assumed by statistical models (Comstock and Robinson, 1948; Robinson et al., 1955; Gouesnard and Gallais, 1992). Seeds for plants designated as male were, however, sown 4 days later than the planting of female parent to allow enough time for a greater range of maturity of female plants mated to a particular male to mature. Also, every plant designated as male was crossed to 4 randomly chosen plants designated as female to avoid assortive mating. Robinson et al. (1955) pointed out that variances by definition, are never negative. Therefore, true values of these negative variances are small positive quantities and negative estimates may have resulted from sampling error. As previously discussed by Comstock and Robinson (1948a, b) and Robinson et al. (1955), estimates of the ratio sigma 2d/sigma 2a presented in Table 3 indicated partial to complete dominance for genes affecting Striga rating 1 and yield of infested plants, although the possibility of overdominance at some loci cannot be ignored. The ratios sigma 2d/sigma 2a of 2.79 and 1.114 obtained for Striga count 1 and yield of non-infested, respectively, suggests that the average degree of dominance for genes affecting these characters were in the complete to overdominance range. Results showed that the proportion of the sigma 2a was greater than that of the sigma 2d (close to zero) for traits like ear Striga rating and anthesis-silk interval of infested plants. Table 3 also indicated additive gene action with partial to complete dominance of genes controlling Striga rating 1 and the yield of infested plants as previously reported by Lonnguist (1953) for maize grain yield. Estimates of genetic correlations. Genotypic and phenotypic correlations between characters are important because they indicate the correlated responses of other characters that may occur during selection of a single trait. Some characters of economic importance like yield, are complex in inheritance and may involve several related characters (Stuber and Moll, 1966). Table 4 shows that yield was negatively correlated with Striga count 1 (rg = -0.22+/-0.46), Striga rating 1 (rg = -0.92+/-0.93) and with ear Striga rating (rg = -0.88+/-1.28). The yield of infested plants was genetically correlated with the number of ear harvested of infested plants (rg = 0.99+/-1.12), and plant height and ear height of infested plants were also, genetically correlated (rg = 0.89+/- 0.52). Similar results were reported by Sturber and Moll (1971) who revealed the presence of genetic interrelationships between yield and number of ears, and plant height and ear height. A low correlation was found between Striga count 1 and Striga rating 1 (rg = 0.20+/-0.41). This indicates that the number of Striga plants is not linearly related to the ability of the host plants to tolerate Striga effects. For instance, a host plant can support few or no Striga plants above ground, but show heavy Striga symptoms. The estimates of genetic correlations between plant height and Striga count 1, Striga rating 1 and ear Striga rating were negative (rg = -0.14+/-0.31, -0.68+/-0.45 and -0.33+/-0.54, respectively). However, genetic correlations between plant height and yield (rg = 0.61+/-0.54) and plant height and ear height (rg = 0.89+/-0.52) showed high and positive values. CONCLUSION The estimates of genetic variances indicated that within the population under study, sigma 2a represents a major portion of the total genotypic variance for characters like ear Striga rating, yield of infested plants, anthesis-silk interval of infested plants and Striga rating. For these traits, additive gene action appears to be the primary source of variation in the population TZLComp 1-C1 and recurrent selection such as mass selection, S1 progeny testing, or half-sib family selection is suggested for improving that population for resistance to Striga. Number of Striga plants is, however, controlled by non-additive gene action which could be interpreted as dominance but, since epistatic variation was not tested, such a distinction could not be drawn. Estimates of genetic correlations indicated that the level of host plant damage was not well correlated with Striga emergence. Thus, resistance to Striga is not controlled by the number of Striga plants attached to the host. To improve the level of Striga resistance and, therefore, the yield of infested plants, it will be more efficient to select for low Striga rating, low ear Striga rating, short anthesis silking intervals, and for low Striga emergence to reduce the bulk of Striga seeds in the soil. ACKNOWLEDGEMENTS This paper is part of a Ph.D thesis submitted by the first author to the University of Ghana, Legon-Accra, Ghana. Academic and Research fundings were provided by IITA/WINROCK fellowship. REFERENCES Anderson, R. L. and Bancroft, T. A. 1952. Statistical Theory in Research. Mc Graw-Hill Book Co., New York. Berner, D. K., Winslow, M. D., Awad, A. E., Cardwell, K. F. and Raj, D. R. M. 1993. Striga Research Methods. A Manual. Draft edition prepared by the IITA Striga research group, pp.71, Ibadan, Nigeria: IITA. Compton, W. A., Gardner, C. O. and Lonnquist, J. H. 1965. Genetic variability in two open-pollinated varieties of corn (Zea mays L.) and their F1 progenies. Crop Science 5:505-508. Comstock, R. E. and Robinson, H. F. 1948a. The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4:254-266. Comstock, R. E. and Robinson, H. F. 1948b. Estimation of average dominance of genes. In: Heterosis. J.W. Gowen (Ed.), pp. 494-516. Iowa State College Press, Ames. Eberhart, S. A., Moll, R. H., Robinson, H. F. and Cockerham, C. C. 1966. Epistatic and other genetic variances in two varieties of maize. Crop Science 6:275-280. Falconer, D. S. 1981. Introduction to Quantitative Genetics. Second edition. Longman, New York. 432pp. Gouesnard, B. and Gallais, A. 1992. Genetic variance component estimation in a nested mating design with positive assortive mating, and application to maize. Crop Science 32:1127-1131. Hallauer, A. R. and Miranda, J. B. 1988. Hereditary Variance: In: Quantitative Genetics in Maize Breeding. Hallauer, A. R., Miranda, J. B. (Eds.), pp. 44-111. Iowa State University Press, Ames. Iowa. Kim, S. K. and Winslow, M. D. 1991. Progress in breeding maize for Striga tolerance/resistance at IITA. In: Proceedings of the 5th International Symposium of Parasitic Weeds. Ransom, J. K., Musselman, L. J., Eorsham, A. D. and Parker, C. (Eds.), pp. 494-499. Nairobi, Kenya. Lindsey, M. F., Lonnguist, J. H. and Gardner, C. O. 1962. Estimates of genetic variance in open-pollinated varieties of cornbelt. Crop Science 5:125-129. Lonnquist, J. H. 1953. Heterosis and yield of grain in maize. Agronomy Journal 45:539-542. Robinson, H. F., Comstock, R.E. and Harvey, P.H. 1955. Genetic variances in open-pollinated varieties of corn. Genetics 40:45-59. SAS Institute. 1987. SAS/STAT Guide for Personal Computers. Version 6.04 ed. SAS Institute, Inc., Cary, NC. Scheinberg, E. 1966. The sampling variance of the correlation coefficients estimated in genetic experiments. Biometrics 22:187-191 Smaling, E. M. A., Stein, A. and Sloot, 1991. A statistical analysis of the influence of Striga hermonthica on maize yields in Kenya. Plant and Soil 138:1-8. Stuber, C. W. and Moll, R. H. 1969. Epistasis in maize (Zea mays L.). 1: F1 hybrids and their S1 progeny. Crop Science 9:124-127. Stuber, C. W. and Moll, R. H. 1971. Epistasis in maize (Zea mays L.). 2: Comparison of selected with unselected populations. Genetics 67:137-149. Copyright 1996 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97001d.gif] [cs97001a.gif] [cs97001c.gif] [cs97001b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}