 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Effect of soybean on subsequent maize grain yield in the guinea savanna zone of West Africa R.J. CARSKY, R. ABAIDOO^1, K. DASHIELL and N. SANGINGA
International Institute of Tropical Agriculture (IITA), PMB 5320, Ibadan,
Nigeria (Received 30 September, 1996; accepted 23 November, 1996)
Code Number: CS97005
Sizes of Files:
Text: 26.6K
Graphics: Tables and line drawings (gif) - 29.2K
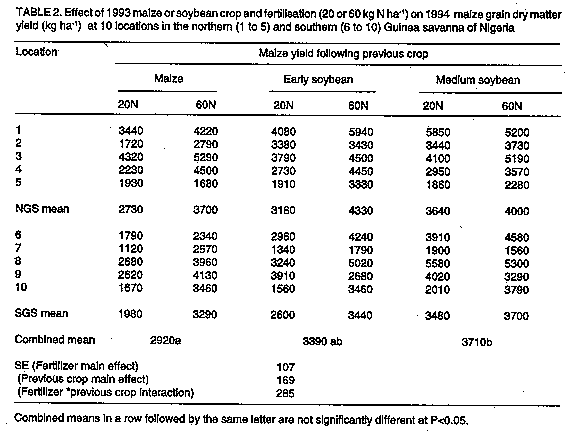
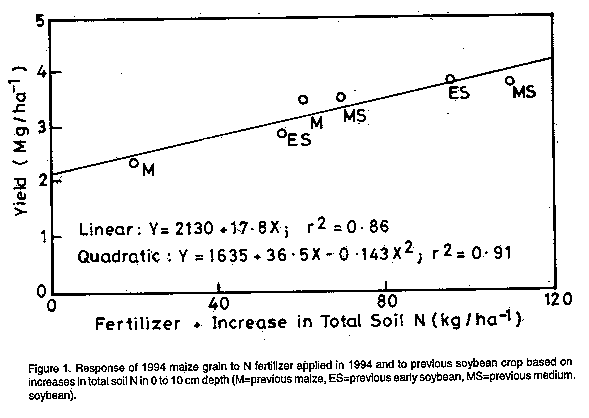
ABSTRACT Two varieties of soybean (Glycine max (L.) Merr.) and a maize (Zea mays L.) control crop were grown in replicated trials at 10 sites in the Guinea savana of Nigeria in 1993 followed by a test crop of maize in 1994 with 20 or 60 kg N ha^-1 to test effect of soybean on subsequent maize grain yield. Maize grain yield increase following soybean was variable but the main effect of previous soybean crop was positive even though the soybean was not inoculated with rhizobia and the aboveground soybean residues, except litter fallen before harvest, were exported from the fields, following the present farmer practice. The yield increase following the medium duration soybean variety (TGx1660-19F) was similar to that from 40 kg N ha^-1 applied 4 weeks after planting to maize preceded by maize. The increase following the early soybean variety (TGx1456-2E) was significantly (P<0.05) smaller than that following the medium duration soybean variety. Total N in the 0 to 10 cm depth of the previous TGx1660-19F plots (0.063%) was significantly greater (P<0.05) than in the previous maize plots (0.058%) when analysed for the combined sites. Thus, the effect of previous soybean crop on maize grain yield was due mostly to residual N availability. Additional research is justified to estimate how soybean crop management influences the yield of subsequent maize in the savanna zone. Key Words: Crop rotation, residue management, N fertilizer RESUME Deux varietes de soya (Glycine max (L) Merr.) et une varietede mas (Zea mays L.) en temps que temoin ont ete cultivees en essais repliques sur 10 sites dans la savane Guineenne du Nigeria en 1993. Ces cultures ont ete suivies d'une culture-test de mais en 1994 avec 20 ou 60 kg N ha^-1. L'augmentation de la production en graine de mais precedee d'une culture de soya par rapport a la production du mais precedee d'une culture de mais etait variable. L'effet principal de la culture de soya etait positif, bien que le soya n'ait pas ete inocule et que le residu du soya aerien, a l'exception du dechet tombe avant la recolte, ait ete exporte des champs a l'example des pratiques courantes. L'augmentation de la recolte de mais precedee de la variete de soja a duree intermediaire (TGx1660-19F) etait semblable a l'augmentation obtenue par application de 40 kg N ha^-1 sous forme d'ureee 4 semaines apres la plantation de mais precede par du mais. L'augmentation suivant la variete de soya precoce (TGx1456-2E) etait significativement plus faible (P < 0.05) que celle suivant la varietede duree intermediaire. L'azote total une profondeur de 0-10cm dans les parcelles de TGx1660-19F (0.063 %) etait significativement plus elevee (P < 0.05) que dans les parcelles temoin (0.058 %) lorsque l'anae est realisee tous sites confondus. Par consequent, l'effet d culture de soya sur la recolte posterieure de mais resulte principalement de la disponibilite residuelle en azote. Des recherches additionnelles sont justifiees pour estimer comment la conduite de la culture du soja influence la recolte posterieure de mais dans la zone de savane Guineenne. Mots Cles: rotation de culture, effet residuelle, equivalence de fertilisation d'azote, credits d'azote INTRODUCTION Soybean (Glycine max L. Merr.) may contribute to the N needs of maize in West Africa. This crop has become increasingly important in Nigeria, and has spread to large parts of the Guinea savanna zone where it is well adapted (Smith et al., 1993a). Although soybean is generally considered to be a crop that depletes soil N (Giller and Cadisch, 1995), this may depend on choice of cultivar and the field history (Peoples et al., 1995b). Soybean may contribute to soil N through biological N2- fixation, some of which can be available to a subsequent maize crop. It may also absorb less soil nitrate than maize, leaving more for a subsequent crop. This "nitrate-sparing" effect may be as important as carry over of biologically fixed N (Peoples et al., 1995a). In any case, nitrogen fertilizer recommendations for maize following soybean might be reduced, making maize production more sustainable from an ecological and economic perspective. Elsewhere in the world, the N fertilizer recommendations for cereal production take previous legume crop into consideration. The N credit from one crop of soybean to the following year's crop grain sorghum in Missouri was 40 to 80 kg ha^-1 depending on soil type (Hanson et al., 1988), in a mechanised system in which soybean residues were returned to the soil. However, in West Africa soybean is harvested by pulling the plant out of the ground and carrying it for threshing; soybean crop residues are rarely returned to the soil. Eaglesham et al. (1982) measured N fixation and seed content of soybean and predicted net depletion of soil N by two varieties of soybean. They did not, however, measure root N content or subsequent effect on a test crop. MacColl (1989) observed an average N fertilizer equivalence of only 7 kg ha^-1 during five years at two sites in Malawi, but soybean was reported to be poorly nodulated. Soybean cultivars very likely influence the contribution of N to the subsequent maize crop. Work with cowpea has shown that the contribution to soil N theoretically increases with the duration of the legume crop as N harvest index decreases (Eaglesham et al., 1982). Stoop and van Stavern (1982) demonstrated that the impact of residual legume N on subsequent millet grain yield increases with increasing maturity cycle of the preceding cowpea. This effect can be hypothesised also for soybean. The purpose of this study was, therefore, to estimate the value of residual soybean N on subsequent maize grain yield under the prevailing situation of soybean residue exportation at 10 sites in the Guinea savanna of northern Nigeria. MATERIALS AND METHODS Five trials each were conducted in northern Guinea savanna (NGS) and southern Guinea savanna (SGS) of Nigeria in northern and southern Kaduna State, respectively. The rainfall pattern is mono-modal and was adequate during the cropping period (June to September) for soybean growth in 1993 (870 mm in NGS and 1030 mm in the SGS). However, during the maize test crop in 1994, rainfall was low in the NGS (700 mm) and excessive in the SGS (1150 mm). Soil properties in the two zones are presented in Table 1. In 1993, soybean and maize crops were established in a randomised complete block design with four replications. Cultivars of soybean were TGx1456-2E (100-105 days to maturity) and TGx1660-19F (115-120 days to maturity). Soybean seed were not inoculated with rhizobia. Spacing for soybean and maize was 0.6 m on 15-20 cm high ridges. Plot size was 2.4 m wide (4 rows per plot) and 6 m long. Aboveground soybean and maize residues, except litter fallen before harvest, were not returned to the plots. Before sowing in 1994, the soil was sampled to 10 cm depth in the ridges of each plot. These samples were air-dried, crushed, and passed through a 0.5-mm seive. They were analysed for total N using a semi-macro Kjeldahl digestion (Bremner, 1965) in 250-ml flasks and an aluminum block digestor. Three 25 to 30 cm high ridges (0.8 m apart) were made in each plot. Basal fertilizer was applied at the rate of 20-8-17 kg N-P-K ha^-1 and incorporated in the ridges. The International Institute of Tropical Agriculture (IITA) maize hybrid 8321-18 was sown early to mid-June and thinned to one stand per 20 cm apart at two weeks after planting (WAP). At 4 WAP, 40 kg N ha^-1 as urea was incorporated into the ridges in two of the four replications at weeding. In the other two replications, no urea was applied. Thus, the two rates of inorganic N were 20 and 60 kg ha^-1. Choice of N rate was constrained by the need to have an acceptable yield on the collaborating farmerselds. A minimum acceptable rate was perceived to be 20 kg ha^-1. A rate which usually gives reasonable returns is 60 kg ha^-1. At physiological maturity, all ears in the central row, excluding the first and last hill (5.6 m length), were harvested in each plot. Grain from the harvested row was weighed after air-drying and shelling early in the dry season. Data on total soil N and maize grain yields were subjected first to analysis of variance individually for each location. For soil N, the treatments were the three previous crops in four replications. For grain yield, the treatment design was a factorial with two N rates and three previous crops, with two replications per location. The 10 locations were then combined for analysis of variance and previous crop means compared for both grain yield and total soil N. RESULTS Maize grain yields across the 10 sites were highly variable (Table 2). For example, yield of maize grain with 20 kg N ha^-1 following previous maize crop ranged from 1.1 to 4.3 t ha^-1. The response to an additional 40 kg N ha^-1 as urea applied 4 weeks after sowing ranged from -240 to +2280 kg ha^-1. Average increase in yield due to application of 40 kg N ha^-1 was 36% in the northern and 67% in the southern sites. Maize grain yields following the soybean treatments were also highly variable. The effect of previous soybean crop on maize grain yield at each site was rarely statistically significant. This could have been due to presence of only two replications per site. At the fertilizer rate of 20 kg N ha^-1, maize yield following medium duration soybean exceeded yield of maize after maize in eight out of 10 locations (Table 2). Average maize grain yield increase following medium duration soybean was 33% in the northern and 76% in the southern sites. The effect of the early soybean variety was not as consistent, exceeding previous maize in seven of 10 locations. Average yield increase due to the previous early soybean was 16% in the northern and 32% in the southern sites. At the 60 kg N ha^-1 fertilizer rate, the effect of previous soybean was less consistent. Maize yield after soybean exceeded maize after maize at six of the 10 sites for the medium duration variety and five of 10 sites for the early variety. In a combined analysis of variance over all sites (Table 3), the main previous crop effect was statistically significant (P<0.05) in spite of high variability. Over both fertilizer rates, maize yield following medium duration soybean was 27% greater than for maize after maize (Table 2). Maize yield after early soybean was 16% greater than after maize but the difference was not statistically significant. The interaction between previous crop treatments and fertilizer rate was weakly significant (P=0.15). This interaction is evident in Table 2 where the response to fertilizer was 1.1 ha^-1 in the previous maize, 1.0 t ha^-1 in the previous early soybean, and 0.3 t ha^-1 in the previous medium soybean crop. The reduced response to inorganic N indicates that N nutrition was less of a constraint to maize following medium duration soybean. The effect of previous crop on total soil N at 0 to 10 cm depth is presented in Table 4 for each site. Total N in the previous TGx1660-19F (medium duration soybean) plots was higher than in the maize plots at all 10 locations, although the differences at each site were rarely statistically significant. In the early soybean variety plots, total soil N was higher than in the previous maize plots at six locations and lower at two locations. In the combined ANOVA over all sites, total soil N in 0 to 10 cm depth was significantly greater (P<0.05) following soybean than following maize. The average difference was 8.5% for the medium soybean and 6.1% for the early soybean (Table 4). Using these values and the bulk density of 1.4 kg dm-3 commonly observed in the zone (Adeoye, 1986; Adeoye and Mohamed-Saleem, 1990), the mean additional total N present in the soil (0-10 cm depth) would be 49 kg ha^-1 following medium soybean and 35 kg ha^-1 following early soybean. Using the estimates of increased total soil N calculated above, a graph was drawn of the maize yield as a function of the rise in total soil N from the previous soybean crop and the 1994 fertilizer N application. The graph (Fig. 1) reveals that, regardless of the treatment maize grain yield follows a linear N response curve which indicates again that N was the major limitation to maize yield in these trials and that the major benefit of the previous soybean crop was due to greater N supply. DISCUSSION Benefits of soybean rotation unrelated to N supply are sometimes evident, such as when the yield of cereal with optimum fertilizer N is less than yield following soybean (Hanson et al., 1988). In the intensified maize cropping systems found in the Nigerian savanna, non-N benefits from soybean may be from reduced nematode and Striga hermonthica parasitism. Weber et al. (1995) observed reduced incidence and severity of nematode damage on maize plants following soybean (TGx923-2E) compared with a previous cereal crop. Striga hermonthica, a serious parasitic weed of maize, can also have it's seed bank reduced by soybean (Parkinson et al., 1987; Alabi et al., 1994). In this set of trials, however, S. hermonthica numbers were low (0 to 2 m-2) in all of the southern locations and one northern location. They ranged from 3 to 10 m-2 in the other northern sites. The effect of previous crop on S. hermonthica parasitism was not observed probably because of variable S. hermonthica densities in the experimental sites. There is evidence showing that N supply was the major influence of soybean on subsequent maize in these trials. Firstly, N was clearly a constraint as shown by the response of maize to a 40 kg N ha^-1 increment. Secondly, yield response to inorganic N was reduced following soybean, especially the medium duration soybean. The N contribution of previous soybean to maize was also inferred from the lack of a response of the subsequent maize crop to N fertilizer by Gallo et al. (1983). Finally, maize yield (Table 2) and total soil N content (Table 4), taken over the ten sites combined, were significantly higher following medium duration soybean than following maize. For both response variables, early soybean had an intermediate effect. The positive effect of previous soybean observed in these trials could have been partially due to previous soybean absorbing less soil N than previous maize, generally known as rate sparing effect (Peoples et al., 1995a). In addition, there was probably additional N fixed and left in the soil for the subsequent maize crop. Unfortunately, N contents of roots and litter of the previous soybean and maize crops were not determined, so the importance of the two effects could not be quantified. In a related pot trial using soil from the same sites, total N fixed (aboveground plus belowground) was about 163 mg pot-1 for TGx1456-2E and 255 mg pot-1 for TGx1990-19F. The latter had much more nodule mass at soybean maturity (R. Abaidoo, unpublished). Thus, the potential difference in N fixation between the two varieties is clear and corresponds to our observed effects on soil N and maize grain yields. One may estimate the amount of N contained in the below-ground and above-ground litter at maturity to try to account for the increase observed in total soil N. The soybean N content in our field study was reliably determined at full bloom. At that time, mean aboveground N content was 27 kg ha^-1 for both varieties in the southern sites. In the northern sites, it was 34 kg ha^-1 for TGx1456-2E and 30 kg ha^-1 for TGx1660-19F. In a related greenhouse study, Abaidoo (unpublished) estimated the increase in above-ground N content between full bloom and maturity to be 170% in TGx1456-2E and 240% in TGx1660-19F. Thus, above-ground N content at maturity can be estimated at approximately 80 kg ha^-1 for TGx1456-2E and 100 kg ha^-1 for TGx1660-19F. Hanway and Weber (1971) found N content in fallen leaves and petioles to be 20 to 30% of total above-ground N in several varieties, years, and fertilizer treatments. The N content of fallen leaves and petioles may, therefore, be estimated at approximately 20 and 25 kg ha^-1 for the two varieties. In the greehouse trial referred to above, Abaidoo (unpublished) estimated N content in roots at maturity to be slightly greater than 10% of total above-ground N content. Total above- and below-ground litter can, therefore, be estimated at 28 kg ha^-1 for TGx1456-2E and 35 kg ha^-1 for TGx1660-19F. Thus, it appears, without the benefit of direct measurement, however, that roots and litter alone may have been responsible for a large part of the increase in total soil N observed, and that soil nitrate sparing by soybean may have accounted for about 20%. Assuming that N supply was the major benefit, N fertilizer equivalence (or N credit) of a previous crop can be estimated (Lory et al., 1995) by comparing maize grain yield following soybean with only 20 kg ha^-1 to that following maize with 20 and 60 kg ha^-1 (Table 2). The maize grain yield with 20 kg N ha^-1 following medium duration soybean is similar to maize after maize with 60 kg N ha^-1. Therefore, N application to maize following medium duration soybean can be reduced by approximately 40 kg ha^-1. Similarly, N fertilisation to maize following early soybean can be reduced by 15 to 20 kg N ha^-1. A more precise estimation of N credits from previous legumes should be done using a full range of N fertilizer levels, which was not possible because of space limitations on collaborating farmerselds. It should be noted that the N fertilizer equivalence value observed in these trials is substantially larger than was reported by MacColl (1989) in Malawi and Bowen et al. (1988). A large N contribution may be achieved by good soybean growth and residue return to the soil, typical of the system studied by Hanson et al. (1988). A small contribution was related to poor nodulation and growth of the soybean in Malawi (MacColl, 1989). It can also be related to the previous control treatment. For example, Bowen et al. (1988) used a bare fallow plot as a control in which 247 kg ha^-1 of inorganic N accumulated in the soil profile compared to 124 kg ha^-1 under soybean. Resulting N uptake (and yield) of maize was similar following both treatments, indicating substantial N supply by soybean residues that was not reflected by the maize test crop. Maize grain yields in the SGS were generally lower than those of NGS (Table 2) although soil N was not significantly different (Table 1). Response to inorganic N following maize was greater in the SGS (67%) than NGS (36%). Response to previous soybean was also greater in the SGS. At the 20 kg N ha^-1 rate, response to previous medium duration soybean was 76% in the SGS and 33% in the NGS. Corresponding values following the early soybean are 32% in the SGS and 16% in the NGS. Lower maize yields in the SGS may have been caused by lower plant available P (Table 1). However, the levels of P were generally in the adequate range. Rather, the cause of lower yields in the SGS is likely to be the sandy soil texture (Table 1) and higher rainfall, which favoured leaching of soil nitrate beyond plant roots. In conclusion, this study estimated the effect of two cultivars of soybean and a maize control on subsequent maize yields. The difference between previous soybean and previous maize was impressive considering that the standing soybean residues were exported from the fields. Although precise recommendations can not be made about choice of soybean cultivar and management, testing of soybean in rotation with maize merits encouragement because the system poses very little economic risks to producers where a market for soybean exists such as in Nigeria. Choice of soybean cultivar, and mangement of the crop and its residues will influence the residual N contribution to subsequent maize. Therefore, future studies should focus on the roles of cultivar characteristics, soybean crop management, and soybean residue management on subsequent maize yields in rotation systems. Estimation of root and litter N contents should be included wherever possible. ACKNOWLEDGEMENTS The trials reported here were manged by A.G. Azeez in the northern Guinea savanna and Rufus Oyom in the southern Guinea savanna. The manuscript was reviewed at IITA by G. Tian, J. Pleysier and I. Kasele. REFERENCES Adeoye, K.B. 1986. Physical changes induced by rainfall in the surface layer of an Alfisol, northern Nigeria. Geoderma 39:59-66. Adeoye, K.B. and Mohamed-Saleem, M.A. 1990. Comparision of effects of some tillage methods on soil physical properties and yield of maize and stylo in a degraded ferruginous tropical soil. Soil and Tillage Research 18:63-72. Alabi, M.O., Berner, D.K. and Olaniyan, G.O. 1994. Characterization of soybean cultivars for Striga hermonthica control. Phyto-pathology 84:1151. Bowen, W.T., Quintana, J.O., Pereira, J., Bouldin, D.R., Reid, W.S. and Lathwell, D.J. 1988. Screening legume green manures as nitrogen source to succeeding non-legume crops. I. The fallow soil method. Plant and Soil 111:75-80. Bremner, J.M. 1965. Total nitrogen. In: Methods of Soil Analysis, Part 2. Black, C.A. (Ed.), pp. 1149-1178. American Society of Agronomy, Madison, Wisconsin. Eaglesham, A.R.J., Ayanaba, A., Ranga Rao, V. and Eskew, D.L. 1982. Mineral N effects on cowpea and soybean crops in a Nigerian soil. II. Amounts of N fixed and accrual to the soil. Plant and Soil 68:183-192. Feller, C. 1995. La matire organique du sol: un indicateur de la fertilite. Application aux zones sahelienne et Soudanienne. Agriculture et Developpement 8:35-41. Gallo, P.B., Sawazaki, E., Hiroce, R. and Mascarenhas, H.A.A. 1983. Producao de miho afetada pelo nitrogenio mineral e cultivos anteriores com soja. Revista Brasileira de Ciencia do Solo 7:149-152. Giller, K.E. and Cadisch, G. 1995. Future benefits from biological nitrogen fixation: An ecological approach to agriculture. Plant and Soil 174:255-277. Hanson, R.G., Stecker, J.A. and Maledy, S.R. 1988. Effect of soybean rotation on the response of sorghum to fertilizer nitrogen. Journal of Production Agriculture 1:318-321. Hanway, J.J. and Weber, C.R. 1971. Accumulation of N, P, and K by soybean (Glycine max (L.) Merrill) plants. Agronomy Journal 63:406-408. Jones, M.J. 1973. The organic matter content of the savanna soils of West Africa. Journal of Soil Science 24:42-53. Lory, J.A., Russelle, M.P. and Peterson, T.A. 1995. A comparison of two nitrogen credit methods: Traditional vs. difference. Agronomy Journal 87:648-651. MacColl, D. 1989. Studies on maize (Zea mays) at Bunda, Malawi. 2. Yield in short rotations with legumes. Experimental Agriculture 25:51-54. Parkinson, V., Efron, Y., Bello, L. and Dashiell, K. 1987. Trap crops as a cultural measure in Striga control in Africa. FAO Plant Protection Bulletin 35:51-54. Peoples, M.B., Herridge, D.F. and Ladha, J.K. 1995a. Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? Plant and Soil 174:3-28. Peoples, M.B., Ladha, J.K. and Herridge, D.F. 1995b. Enhancing legume N2 fixation through plant and soil management. Plant and Soil 174:83-101. Smith, J., Woodworth, J.B. and Dashiell, K.E. 1993a. Government policy and farm level technologies: The expansion of soybean in Nigeria. Agricultural Systems in Africa 3:20-32. Smith, J., Barau, A.D., Goldman, A. and Mareck, J.H. 1993b. The role of technology in agricultural intensification: The evolution of maize production in the northern Guinea savannah of Nigeria. In: Social Science Research for Agricultural Technology Development: Spatial and Temporal Dimensions. Dvork, K.A. (Ed.), pp. 144-166. CAB International, Wallingford, UK. Stoop, W.A. and van Staveren, J.P. 1982. Effect of cowpeas in cereal rotations on subsequent crop yields under semi-arid conditions in Upper Volta. In: Biological Nitrogen Fixation Technology for Tropical Agriculture. Graham, P.H. and Harris, S.C. (Eds.), pp. 653-657. CIAT, Cali, Colombia. Weber, G.K., Chindo, P.S., Elemo, K.A. and Oikeh, S. 1995. Nematodes as Production Constraints in Intensifying Cereal-Based Cropping Systems of the Northern Guinea Savanna. Resource and Crop Management Research Monograph No. 17. IITA,Ibadan, Nigeria. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97005c.gif] [cs97005a.gif] [cs97005b.gif] [cs97005e.gif] [cs97005d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}