 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Feasibility of chemical control of the African bollworm on sunflower in South Africa H. DU PLESSIS Grain Crops Institute, Agricultural Research Council, Private Bag X1251, Potchefstroom, 2520, Republic of South Africa (Recieved 11 September, 1996; accepted 31 December, 1996 )
Code Number: CS97007
Sizes of Files:
Text: 19.1K
Graphics: Tables and line drawings (gif) - 23.1K
ABSTRACTThe efficacy of methomyl and fenvalerate as a tank mixture for the control of African bollworm (Helicoverpa armigera Hubner) was compared by means of aerial application from two directions in relation to row direction and two plant reproductive stages (budding R-3, and during flowering and pollination, R-5) of sunflower (Helianthus annuus L.). Chemical control was effective during both reproductive stages, reductions in pest numbers ranged from 55.3% during R-3, to 86.5% during R-5, irrespective of the direction of application. Cost of control constituted 14% of the net profit margin. To determine the effect on yield and the ability of sunflower to compensate, insect damage was simulated on florets before pollination as well as on young achenes in the milky stage. A reduction in yield, although not significantly different from the undamaged control heads, was observed at damage level greater than 20%. Due to the cost of aerial insecticide application and the ability of sunflower to compensate for losses, chemical control of H. armigera should not be applied at first signs of eggs or larvae. Based on observations in this study, it is proposed that insecticide application be done when at least 20% damage per head occurs. Key Words: Helicoverpa armigera, Lepidoptera, compensation, Helianthus annuus RESUME L'efficacite de methomyl et de fenvalerate pour controler le ver du grain Africain (Helicoverpa armigera Hubner) etait comparee par une application aerienne en deux directions en relation avec la direction en rangees, et a deux phases reproductives, (phase bouton, R-3 et pendant la floraison, et phase pollinisation, R-5) du tournesol (Helianthus annuus L.). Le controle chimique etait efficace pendant les deux phases reproductives et la reduction du nombre d'insectes variait entre 55.3 % en R-3, a 86.5 % en R-5, independamment de la direction d'application. Les couts du controle constituaient 14 % de la marge beneficiaire nette. Pour determiner l'effet sur la recolte et la possibilite de compensation du tournesol, l' attaque des insectes etait simulee sur des fleurons avant pollinisation et aussi sur les akenes a la phase laiteuse. Une reduction de la recolte, mais pas significativement differente des temoins, etait observee a un niveau d'attaque de plus que 20 %. Comme les insecticides sont chers et le tournesol possede la capacite de compenser des pertes, un controle chimique n'est pas recommande des qu'on observe des oeufs ou des larves. Se basant sur les observations de cette etude, on propose une application d'insecticide quand 20 % d'attaque par tete de fleur apparait. Mots Cles: le ver du grain Africain, controle chimique, compensation, Helianthus annuus INTRODUCTION African bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae), is regularly present during the reproductive stage of cultivated sunflower (Helianthus annuus L.) in South Africa. However, little reseach on H. armigera, as a pest of sunflower, has been conducted in this country (Von Maltitz, 1993). Levels of infestation vary between localities and seasons, sporadically reaching epidemic proportions. No economic injury level as a guideline for control exists, and many producers adhere to the current chemical control recommendation which is to apply insecticides as soon as eggs or young larvae are noticed (Krause et al., 1996). All currently registered insecticides for control of H. armigera on sunflower have a contact action and are aerially applied (Krause et al., 1996). Larvae of all instars burrow under and between bracts into the receptacle (Von Maltitz, 1993), and are, therefore, often not exposed to insecticides. Effective insecticide application is also limited by the downward inclination of sunflower heads or by the fact that heads are often turned in the opposite direction to aerial application. In addition to the uncertainty concerning the efficacy of chemical control, data is needed on the economic importance of various larval stages, damage levels, susceptability of various plant growth stages and the possibility of yield loss compensation by plants attacked by the African bollworm.
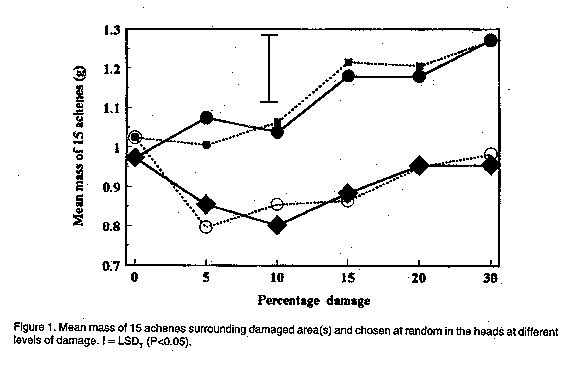
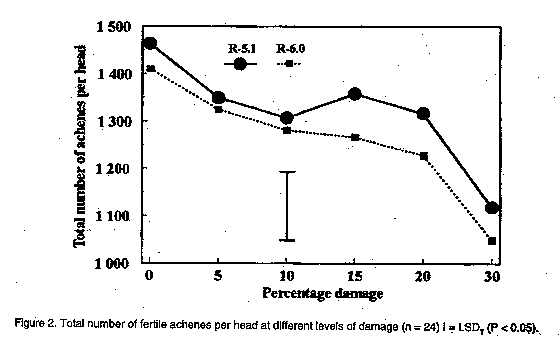
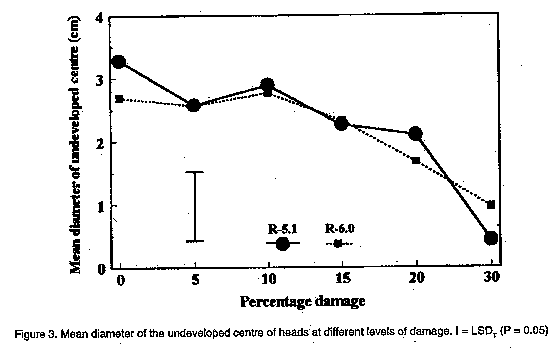
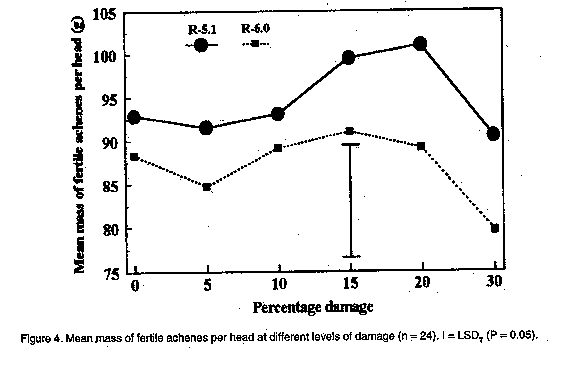
In the present investigation, the efficacy of insecticides applied to sunflower was compared by means of aerial application from two directions at two reproductive stages of sunflower. The effect of simulated insect damage to florets before pollination, as well as to young achenes in the milky stage, on yield, and the ability of sunflower to compensate for damage was also determined. MATERIALS AND METHODS Efficacy of aerial application. The sunflower cultivar PAN 7392 was planted in mid-January 1995 in a commercial field in the Bothaville district (27 degrees 23'S, 26 degrees 37'E). The distance between rows was 90 cm with an inter-plant spacing of 40 cm, equivalent to 28,000 plants ha^-1. Treatments involved evaluation of efficacy of methomyl and fenvalerate on H. armigera, as well as the possible effects of direction of insecticide application and plant growth stage on insecticide efficacy. Methomyl (90 g a.i. ha^-1) and fenvalerate (20 g a.i. ha^-1) were applied aerially as a tank mixture in 30 l of water ha^-1 to a section in the centre of the field. Swath width was 23 m. Two sub-treatments consisted of the direction in which sunflower heads were facing in relation to the direction of application, viz. to the back (posterior) and to the front of the head (anterior). Two reproductive plant stages were evaluated, viz. R-3 (budding stage) and R-5 (anthesis and pollination). Two data rows in the centre of one swath for each of the two directions of application were selected before application . Two untreated check rows were selected from the unsprayed section of the field, five rows apart and 46 m from the treated area to avoid insecticide drift effects. During the pre-spray assessment, bract leaves of R-3 heads were opened and the number of larvae on and inside buds recorded. For R-5, larval numbers were recorded for both surfaces of the heads, as well as those underneath bract leaves. Twenty heads of R-5 from each treatment row (40 in total) and the untreated control were selected and tagged, based on the presence of larvae before application. Since immature buds are not heliotropic, R-3 applied to only one direction of application. Due to the presence of a few dead larvae (apparently from a polyhedrosis virus), post-application assessments were done in quick succession, viz. eight hours and five days after application. Efficacy of control was calculated according to the Behrens-Fischer t-test and Henderson and Tilton method (Henderson and Tilton, 1955), where survival of larvae on control plants were taken into account. Yield was determined at harvest. Yield loss compensation. A field trial was conducted at Potchefstroom (26 degrees 43'S, 27degrees 06'E), during the 1994/95 season. The sunflower hybrid, SNK 43 (75% self-compatible), was planted in mid-December in 15 rows, 40 m in length, with 90 cm between rows and 40 cm inter-plant spacing. Plants were thinned two weeks after emergence to ensure an even plant stand. Irrigation was provided as needed. No natural H. armigera infestation occurred and insect control was, therefore, not applied. Treatments were applied at two reproductive stages, viz. R-5.1 and R-6.0. R-5.1 is the stage at which 10% of the head area (disk flowers) has completed or is in flower. R-6.0 represents the reproductive stage in which anthesis and pollination are completed and the ray flowers are wilting (Schneiter and Miller, 1981). During each reproductive stage, florets or achenes were removed by hand to simulate insect damage. Plants were selected from the central 13 rows, allowing one guard row at each edge. In a preliminary assessment during each of the two reproductive stages, the average diameter of 30 heads was determined. This was done by making two perpendicular measurements across the head in order to homogenise variation in head size within a specific reproductive stage (R-5.1 = 10 cm and R-6.0 = 20 cm). From these, the percentage area of florets or achenes to be removed from heads was calculated. Six treatments were used, involving the removal of 0, 5, 10, 15, 20 and 30% florets or achenes. Each treatment had 24 replications (heads). A circular area of florets or achenes was removed with a pinset in increments of 5% for 5, 10 and 15% damage, and areas in increments of 10% for 20 and 30% damage. The centres of heads which are often undeveloped were not used. Since only heads with the predetermined diameter were used, randomisation was automatically obtained. To avoid any external damage to achenes by birds and to ensure maximum pollination, all heads were covered with gauze bags at the R-6.0 growth stage. Heads were harvested and dried after physiological maturity. The average diameter of heads and undeveloped centres were determined by means of two perpendicular measurements across each head and centre, respectively. The number and mass of fertile achenes were determined for each head. Additionally, the mass of 15 achenes surrounding each of the damaged areas in a head, as well as that of 15 randomly selected achenes per damaged and undamaged head were determined.
The data were subjected to factorial analyses using plant growth stage as main factor and injury levels as sub-factor. To eliminate differences in head diameter after maturity, head diameter was used as a covariant in analyses of both total number and total mass of achenes per head. RESULTS AND DISCUSSION Efficacy of control. Larvae occurred from the budding stage onwards, which confirms the results of Von Maltitz (1993). First and second instar larvae were mainly found on the sunflower buds, with a feeding preference for involucral bracts. Later instar larvae fed more extensively than younger larvae, consequently doing more damage. One mature larva feeding inside a bud may completely destroy the immature florets. However, plants infested at the budding stage escaped serious feeding damage as the majority of larvae matured during anthesis, which is in accordance with results by Von Maltitz (1993).
The mean number of larvae found on heads during R-5 was 4.25, with a maximum of 12. According to Von Maltitz (1993), from larvae occurring on sunflower, the percentage feeding on achenes increases from zero for the first instar to 6% for the final instar. Damage caused to achenes during this study was slight, which supports results by Broadley and Ironside (1980) which suggest that sunflower plants are able to support a number of larvae without a significant effect on yield.
The results of aerial application in this study are provided in Table 1. Chemical control was effective during the budding stage (R-3) and anthesis (R-5), irrespective of the direction of application. Taking the pre-spray counts into consideration, the efficacy calculated according to the formula by Henderson and Tilton (Henderson and Tilton, 1955) was 32.9% and 55.3% on R-3 stage plants for counts done eight hours and five days after application, respectively. Application directed at the posterior side of heads in the R-5 stage was 50.4% and 86.5% effective at eight hours and five days, respectively, while anterior application was 39.5% and 84.2% effective on the two post spray counts, respectively. The total cost of aerial application was R 59.40 ha^-1, of which R 29.40 ha^-1 accounted for direct cost of insecticide. Yield was 1.2 t ha^-1 . With the current producer price of R 950 t-1 for sunflower, the resulting gross income was R 1,140 ha^-1, with a net profit margin of R 450 t-1. The cost of control, therefore, constituted 13.81% of the net profit margin, indicating that a loss of approximately 14% may justify chemical control. However, damage, and concomitant yield loss, cannot be related to larval numbers, since various instars consumed different amounts of plant tissue. Furthermore, a definite preference for burrowing under the bract leaves into the pith tissue of the receptacle, which would not necessarily result in yield loss, was observed. It, therefore, appears that actual damage to achenes, rather than the number of larvae, should be taken into account when considering economic injury levels. Yield loss compensation. The mean mass of 15 achenes surrounding damaged areas was less than the mean mass of 15 randomly chosen achenes. This applied to both reproductive stages (Fig. 1), indicating that achenes surrounding damaged areas do not compensate for loss of florets and achenes. However, the mean mass of randomly chosen achenes increased at increased levels of damage. Compensation for lost florets and achenes, therefore, occurred over the entire head. Compared with undamaged heads, the mean mass of achenes was significantly higher at levels of 15, 20 and 30% damage at both reproductive stages. The total number of achenes produced by undamaged heads differed significantly from those with 10 and 30% levels of damage at the R-5.1 stage (Fig. 2). The reduced number of achenes at a 10% damage level compared with levels of 15 and 20% damage (R-5.1) may be explained by an increase in the undeveloped centre area of heads at a 10% level of damage (Fig. 3). In general, the total number of fertile achenes produced per head damaged at R-6.0, decreased with increasing levels of damage (Fig. 2). The significant difference observed in the number of achenes produced by damaged and undamaged heads may be attributed to the exceptionally large proportion of the head involved at damage levels above 20%. Although the size of undeveloped centre areas decreased at increased levels of damage (Fig. 3), this was apparently unable to compensate for loss of achenes. No significant differences (P < 0.05) in the total mass of fertile achenes per head were observed between damaged and undamaged heads, up to a 30% level of damage (Fig. 4). This applied to both reproductive stages, and may be attributed to the increase in achene mass over the entire head as well as to a reduction in size of undeveloped centres. These results differ from those of Charlet and Miller (1993), who reported that the increase in mass of individual achenes was not sufficient to compensate for loss of florets before pollination at a level of only 10% damage. Sedgewick et al. (1986) found sunflower to compensate for up to 15% damage during the soft dough stage compared to 30% found during this study. The difference may be related to factors such as sunflower variety and climatic effects. CONCLUSION Actual damage is the only criterion which can be used in the determination of economic injury levels for control of African bollworm on sunflower. Aerial application of insecticides was effective during both the R-3 and R-5 growth stages, when later instars, which are more damaging, occurred on the heads. According to the net profit margin, a damage level of 14% to florets and achenes justified chemical control. However, due to preferential feeding on sites other than achenes, as well as to the ability of plants to compensate for damage to florets and achenes, a significant number of larvae can be tolerated without any significant effect on yield. Due to the high cost involved in aerial application of agrochemicals and the ability of sunflower to compensate for losses, chemical control of H. armigera should not be applied at first signs of eggs and larvae as indicated by Krause et al. (1996). A reduction in yield, although not significantly different from undamaged control heads was observed at a damage level greater than 20%. It is proposed that insecticides be applied when at least 20% damage per head occurs, in order to promote timely control measures. ACKNOWLEDGEMENTS Messrs D. Dreyer and T. Rossouw for permission to work on their farms, Mrs U. du Plessis (ARC - Grain Crops Institute) and J. Bothma (Sanachem) for technical assistance, PJ van Rooyen for statistical assistance and Dr J.B.J. van Rensburg (ARC - Grain Crops Instititute) for critical reading of the manuscript. REFERENCES Broadley, R.H. and Ironside, D.A. 1980. Insect pests of sunflower. Part 1. Queensland Agricultural Science Journal 106:xxv-xxviii. Charlet, L.D. and Miller, J.F. 1993. Seed production after floret removal from sunflower heads. Agronomy Journal 85:56-58. Henderson, C.F. and Tilton, E.W. 1955. Tests with acaricides against the brown wheat mite. Journal of Economic Entomology 48:157-161. Krause, M., Nel, A. and Van Zyl, K. 1996. A Guide to the Use of Pesticides and Fungicides in the Republic of South Africa. Government Printer, Pretoria. 183pp. Schneiter, A.A. and Miller, J.F. 1981. Stages of sunflower development. Crop Science 21:901-903.
Sedgewick, J.A., Oldemeyer, J.L. and Swenson, E.L. 1986. Shrinkage and growth compensation in common sunflowers: Refining estimates of damage. Journal of Wildlife Management 50:513-520. Von Maltitz, E. F. 1993. The American bollworm, Heliothis armigera (Hubner) (Lepidoptera: Noctuidae) on sunflower. II. Feeding site preferences of the larvae. Phytophylactica 25:249-252. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97007e.gif] [cs97007b.gif] [cs97007c.gif] [cs97007a.gif] [cs97007d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}