 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
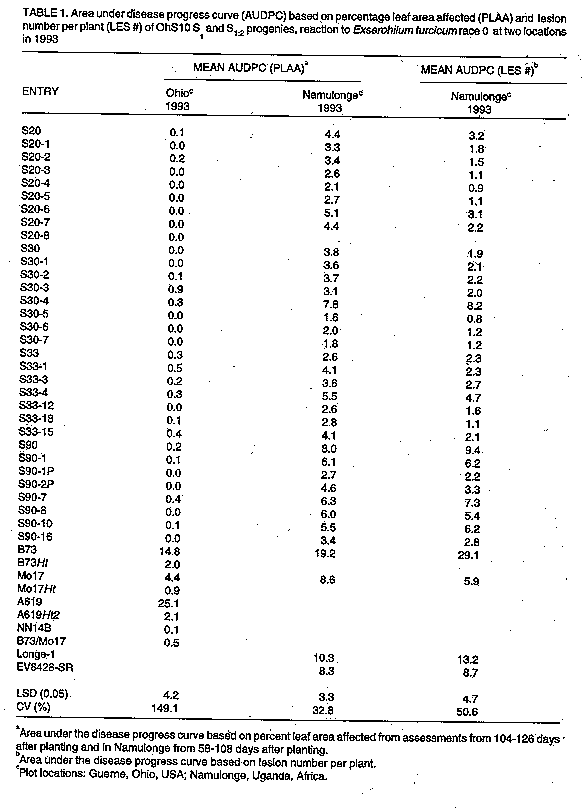
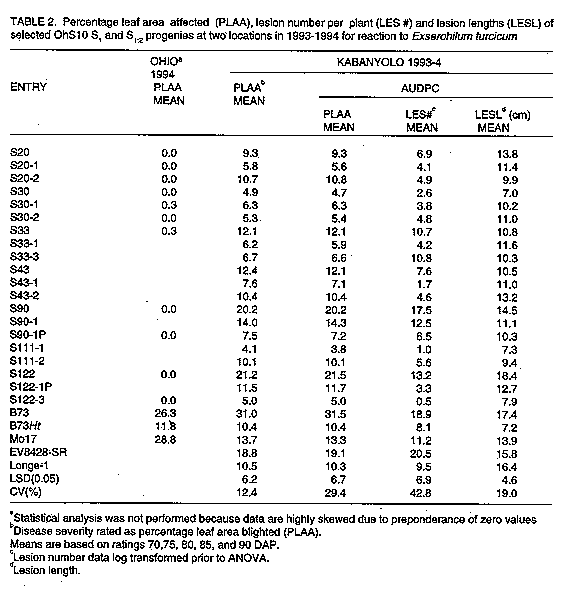
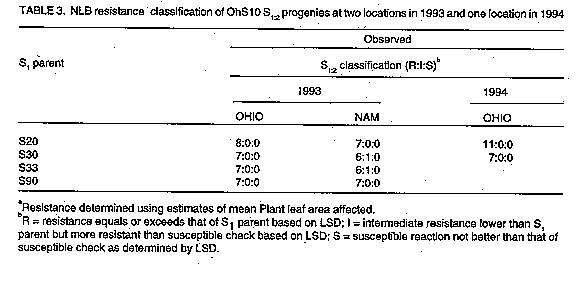
Inheritance of race-nonspecific resistance to Exserohilum turcicum in maize synthetic population OhS10 R.C. PRATT, P.E. LIPPS^1, F. SSANGO^2+, J.J. HAKIZA^3 and E. ADIPALA^2
Department of Horticulture and Crop Science, Ohio Agricultural Research and
Development Center,Wooster, Ohio, U.S.A. 44691 (Received 4 July, 1996; accepted 21 February, 1997)
Code Number: CS97008
Sizes of Files:
Text: 27.5K
Graphics: Tables (gif) - 27.3K
ABSTRACT Progeny tests of resistant lines derived from maize (Zea mays L.) synthetic population OhS10 were conducted at one location during two years (1993-94) in Ohio (Guerne) and at two locations during one year (1993) in Uganda (Kabanyolo and Namulonge) to examine the inheritance of resistance to Exserohilum turcicum(Pass.), causal agent of northern leaf blight (NLB). Individual plants in single ear derived selfed progenies (S1 and S1:2) were inoculated in a controlled fashion (Guerne, Namulonge) or naturally infected (Kabanyolo). Host responses to infection were characterized by determining area under the disease progress curve (AUDPC), calculated from percent leaf area affected (PLAA) and lesion number counts, and lesion size. The majority of S1 and S1:2 progenies had few lesions, low rates of disease development, and high levels of resistance, indicating that multiple factors govern expression of resistance. Of the progenies tested, S20-4, S30-5, S30-6 and S30-7 exhibited consistently high levels of resistance to NLB. Key Words: Breeding, maize, genetics, host resistance RESUME Des tests de descendance des lignees resistantes obtenues a partir d'une population synthetique de mais OhS10 ont ete executes a un endroit pendant deux annees (1993-1994) Ohio (Guerne) et a deux endroits pendant une annee (1993) en Ouganda (Kabanyolo et Namulonge) pour examiner la transmission de la resistance a Exserohilum turcicum (Pass.), qui cause le rouille des feuilles (qui sevit au nord) (NLB). Les plantes individuelles des progenies obtenues par descendance autofecondee issue d'un epis unique (S1 et S1:2) etaient inoculees de facon controee (Guerne, Namulonge) ou infectees naturellement (Kabanyolo). Les reponses des infections etaient caracterisees par la determination de la surface en dessous de la courbe de progres de la maladie (AUDPC), qui etaient calculee a partir du pourcentage de surface de la feuille affectee (PLAA) et le nombre des lesions et la taille de lesions. La majoritedes descendants S1 et S1:2 avaient peu de lesions, un developpement ralentie de la maladie et un niveau de resistance eleve, ce qui indique que plusieurs paramtres influencent l'expression de la resistance. S20-4, S30-5, S30-6 et S30-7 montrent un niveau de resistance eleve contre NLB. Mots Cles: amelioration, le mais, la genetique, resistance INTRODUCTION Exserohilum turcicum (Pass.) Leonard & Suggs, causal agent of northern leaf blight (NLB), is an important fungal pathogen of maize (Zea mays L.) in many agricultural regions (Ullstrup, 1977; Ceballos et al., 1991). When NLB epidemics begin before silking, and environmental conditions favour disease development, yield losses may exceed 40% (Ullstrup and Miles, 1957; Raymundo and Hooker, 1981; Perkins and Pedersen, 1987). Both race-specific and race-nonspecific host resistance to E. turcicum are available to maize breeders (Hughes and Hooker, 1971; Meyer et al., 1991; Freymark et al., 1993). Monogenic, race-specific resistance, under the control of Ht1, Ht2, and Ht3 genes, is expressed as chlorotic-lesion type resistance (Hooker, 1961; Hooker, 1967). HtN based resistance is expressed as delayed lesion development in adult plants (Gevers, 1975). Another major gene, designated HtM, has recently been described (Robbins and Warren, 1993). Race-nonspecific resistance is typically characterised by fewer lesions and a lower rate of disease development than observed in susceptible genotypes (Hughes and Hooker, 1971; Hooker and Kim, 1973; Sigulas et al., 1988; Adipala et al., 1993a). Race-nonspecific resistance is desirable because it is effective against all physiological races of E. turcicum (Hooker and Kim, 1973). Single gene resistance is vulnerable to the development of new races (Turner and Johnson, 1980) whereas polygenic resistance is not easily overcome by new races (Johnson and Jellis, 1992). Race-nonspecific, rate-reducing resistance is frequently considered, even in absence of genetic evidence, to be inherited polygenically (Jenkins and Robert, 1952; Jenkins, 1957; Hughes and Hooker, 1971; Hooker and Kim, 1973). A normal distribution of resistance in an F2 population derived from a cross between resistant and susceptible parents is likely to be the result of polygenic inheritance. Other interpretations are possible due to environment influences and genotype-environment interaction. A single gene may also effect what appears to be a rate-reducing response (reduction in "r") characteristic of several genes (Martin and Ellingboe, 1975), rather than a qualitative response. Indeed. the Ht genes do result in a marked reduction in sporulation, a component of rate-reducing resistance. Researchers have traditionally verified rate-reducing resistance using area under the disease progress curve (AUDPC) values, calculated with multiple ratings of percent leaf area affected (PLAA) (Ceballos et al., 1991). Rate-reducing resistance of inbred Mo 17 to NLB is governed by multiple quantitative trait loci (Freymark et al., 1993; Dingerdissen et al., 1996). Many improved open-pollinated cultivars of maize with potential for cultivation in Uganda are susceptible to NLB (Adipala et al., 1993b). Several of the open-pollinated cultivars and germplasm populations display race-nonspecific resistance to races 0 and 1 and rate-reducing resistance responses (Adipala et al., 1993a; Pratt et al., 1993). The present emphasis on development of cultivars with polygenic, race-nonspecific resistance, instead of monogenic, race-specific resistance, is intended to promote the longevity of resistance factors and suppress formation of new races of NLB. One study identified race 0 in Uganda (Adipala et al., 1993b) and another suggested the presence of multiple races (Welz et al., 1993). Comprehensive genetic studies to confirm polygenic inheritance of resistance in potential donor populations are problematic. Populations with a broad genetic base are often comprised of both resistant and susceptible genotypes (Adipala et al., 1993a; Pratt et al., 1993), with resistance genes segregating and allelic frequencies unknown. Successful identification and transfer of desirable alleles from these populations requires isolation of resistant individuals or progenies, characterising the resistance, and using effective selection procedures to transfer resistance genes to an acceptable agronomic genotype. The objective of this study was to determine if race-nonspecific resistance in partially inbred lines from a synthetic population (OhS10) was attributable to multiple resistance factors. We made these determinations with a family structure (S1-S2) frequently employed in both population improvement and hybrid development breeding programmes. B73Ht, Mo 17Ht, NN 14B) were also included in the Ohio experiment to provide visual checks for chlorotic lesion symptoms. MATERIALS AND METHODS Host plants. The OhS10 synthetic population is comprised of one-half tropical maize germplasm and one-half United States Corn Belt germplasm. It is a potential resource to maize breeders in both temperate and tropical regions. Development of OhS10 was initiated by William R. Findley (USDA/ARS, retired) by crossing OhS7 by OhS8. It was produced using 14 U.S. Corn Belt inbreds and two tropical races of maize, Tuxpeno and Cuban Flint (Pratt et al. 1994). One hundred and forty-nine progenies were evaluated and some of the most resistant were reserved for study (Pratt et al., 1993). Controlled self-pollination of plants within seven resistant S1 lines (S20, S30, S33, S43, S90, S111, S122) were subsequently undertaken to provide multiple, single ear derived progenies (S 1:2). Experimental design. We assumed that highly resistant S1 lines could be either uniformly resistant (derived from homozygous resistant S0 plants with a single resistance gene, R) or segregating 1:2:1 (RR:Rr:rr), or polygenic resistant. We hypothesised that if resistance was monogenic, examination of individual plants in resistant S1 lines would reveal those lines derived from an Rr S0 plant segregating 3 resistant: 1 susceptible. We also chose to ascertain the genotype of S1 plants by conducting S 1:2 progeny tests. If resistance was polygenically controlled, examination of progenies might reveal some variation in degree of resistance, but no progenies would be expected to display monogenic chlorotic lesion responses, nor would they segregate in a 3:1 manner. For comparison, maize inbreds B73 (susceptible check) and Mo 17 (race non-specific, polygenic resistant check) were included in all experiments. Additionally, susceptible (EV8428SR), intermediate or moderately resistant (Longel) open-pollinated cultivars (Adipala et al., 1993a) were included in experiments conducted in Uganda and susceptible check A619 was included in Ohio experiments. Several Ht and Ht2 stocks (A619Ht2, B73Ht, Mo17Ht, NN14B) were also included in the Ohio experiment to provide visual checks for chlorotic lesion symptoms. Field plots. Plots were established at the Ohio Agricultural Research and Development Center near Guerne, OH, USA, at the Makerere University Agricultural Research Institute at Kabanyolo, Uganda, and at the Namulonge Agricultural and Animal Production Research Institute, Uganda. Throughout the rest of this paper, experiments will be identified by location and year as appropriate. In the Gurne (1993) and Namulonge studies, kernels of 4 S1 lines, 7-8 S 1:2 progenies representing each of the 4 S1 lines, and checks, were planted on 14 May 1993, and on 27 March 1993, respectively, in single row-plots in a randomised complete block design with two replicates. Six resistant S1 lines (S20, S30, S33, S43, S90, S122) and 2 S 1:2 progenies for each S1 line were planted at Kabanyolo on 5 May 1993 in single-row plots in a randomised complete block design with two replicates. Replicates at each location were 3 m long. In Uganda, replicates contained approximately 12 plants and in Ohio approximately 15 plants. In the Guerne (1994) plot, in order to increase spread of E. turcicum, kernels of B37 (Guerne 1993) or B73 x B37 (Guerne 1994) or Kawanda Composite (Namulonge and Kabanyolo, 1993) were planted as two or four-row borders around the entire experimental plot. Plots in Ohio had been maintained in an oat-wheat-maize rotation and were fall ploughed. Manure-bedding straw was applied to the Guerne 1994 field plot and chisel ploughed the following spring. The plots were fertilised with 448 kg ammonium nitrate (33-0-0) and 112 kg (0-18-36) per hectare, incorporated by chisel ploughing. Sutan plus and Bladex 4L herbicides were applied pre-plant at 4.17 and 4.64 1 ha^-1, respectively. Plots were hand-planted on 31 May 1994. In Uganda, plots were established in fields previously planted to soybean [Glycine max (L.) Merr.] (Namulonge) or maize (Kabanyolo). No fertilizers or herbicides were applied, and plots were subsequently hand-weeded when the average maturity of progenies was at growth stages (GS) V1 (first true leaf collar visible), V6 (sixth true leaf collar visible), V12 (twelfth true collar leaf visible), V18 (eighteenth true leaf collar visible), and R2 ("blister" stage, 10-14 days after silking) (Ritchie et al., 1989). Races and inoculum preparation. Monoconidial cultures of field isolates of race 0 from Guerne, Ohio, and race 0 from Namulonge, Uganda, were grown on Difco Potato Dextrose Agar (PDA) (Difco Laboratories, Detroit, MI, USA) in Ohio and Uganda, respectively. After 12-14 days at 21-23 C under fluorescent lights for 1 hr day^-1, colonised agar cultures were removed from petri dishes and placed on autoclaved oat (Avena sativa L.) kernels (Ohio) or sorghum [Sorghum biclor (L.) Moench] seeds (Uganda) in 2-l flasks. Flasks were shaken once daily. After 10-14 days, infested seeds/kernels were air-dried for 2-4 days and stored at 6 C. Inoculations. Entry and spreader plants were inoculated 70 days after planting (DAP) at Namulonge with the Namulonge isolate, by placing approximately 5 g infested sorghum seeds into the whorl of each plant. In Ohio (1993), plants were inoculated 61 DAP with oat kernels, but were inoculated 51 and 58 DAP in 1994. No artificial inoculation was performed at Kabanyolo because NLB infested the maize crop during the previous season and we assumed that natural disease would be high enough to show differential reactions. In Ohio, after each inoculation, 2.5 cm of water was applied for 12 hr overnight using overhead sprinkler irrigation. Data collection. Disease severity was assessed using a rating scale of 0, 0.5, 1, 5, 10, 25, 50, and >75% plant leaf area affected (PLAA). Five PLAA determinations were made at five day intervals at Kabanyolo beginning 70 DAP and continuing until 90 DAP. Four ratings were made at Namulonge using the same scale on four rating dates during the period 58-108 DAP. In Ohio (1993), disease was rated at one weekly intervals beginning at 101 DAP and terminating at 126 DAP, and in 1994 two ratings were taken at 107 DAP and 117 DAP. At Kabanyolo and Namulonge, the mean number of lesions per plant was determined from four counts beginning at 51 DAP and continuing until 72 DAP at Kabanyolo and 58-108 DAP at Namulonge. Lesion length (cm) was measured once at 90 DAP at Kabanyolo. A total of 10-20 lesions for each progeny plot were measured. Measurements were made on an individual plant basis (approximately 24 plants/genotype) at Kabanyolo, and on a plot average basis at Namulonge and in Ohio. Data analysis. Based on multiple assessments of PLAA and lesion number counts, AUDPC values were calculated using the midpoint rule standardised by dividing AUDPC values by the number of days from the first to last assessment for each observation plot (Campbell and Madden, 1990). Analyses of disease severity assessment data were conducted within locations using SAS PROC GLM (SAS, 1985) to determine differences among OhS10 progenies. All mean comparisons were conducted at P<0.05 probability level using Fisher's Proteted Least Significant Difference (LSD) (Steel and Torrie, 1980). RESULTS Environmental conditions favoured development of NLB in Uganda. Late season PLAA ratings at Namulonge ranged from 3 to 28%. At Kabanyolo late season PLAA values ranged from 7 to 45%. In Ohio (1993) late season disease severity ranged from 0 to 1% on OhS10 S1 and S1:2 progenies to 48% on A619. Despite a delayed planting and an additional inoculation, all OhS10 progenies had less than 1% leaf area affected in 1994. Disease did not develop on the majority of progenies in Ohio, but disease severity of A619 reached 65% in 1994 compared to 48% in 1993. The higher disease severity in Uganda permitted better seperation of progenies based on their host responses to NLB. Segregation of susceptible plants was not observed in six S1 lines at Kabanyolo. In both Ohio (1993-4) and Uganda, few significant differences in resistance of sibling S1:2 progenies were detected (Table 1 and table 2). Only two progenies (S30-4 and S33-18) were significantly more susceptible than their S1 progenitors, but neither was as susceptible as the susceptible check B73. Two progenies (S90-1P and S90-16) were more resistant than their S1 parents. The frequency of these differences (2/26) is only slightly more than the frequency expected due to chance alone (1/20). We planted some of the same progenies in 1994 as in 1993 and again B73 showed disease development in Ohio, but little disease progress was evident in the OhS10 progenies (Table 2). A total of eleven S2 progenies of S20 were tested and none of them were susceptible (range 0-1% PLAA). Number of lesion counts indicated that all OhS10 progenies, except S90, had fewer lesions than the susceptible check B73 (Table 2). All S1:2 progenies had fewer lesions than B73 and the majority had fewer than their S1 progenitors. Lack of S1:2 progenies segregating for a fully susceptible phenotype precluded Chi-square goodness of fit tests to examine a 3:1 segregation ratio. DISCUSSION From these data we inferred that resistance in OhS10 S1 lines examined was not controlled by a single gene (Table 3). The high level of resistance observed in selected S1 lines, and the low lesion numbers of resistant S1:2 progenies (equal to or lower than that of Mo17) in earlier studies (Pratt et al., 1993) pointed to a race-nonspecific, likely polygenic, basis of resistance (Windes and Pedersen, 1991). Because both resistant and susceptible genotypes were included in the OhS10 population, and because no parents were known to have single-gene (Ht) resistance, we assumed that highly resistant S1 lines contained multiple resistance factors. We also assumed that individuals within S1 lines possessed resistance factors at multiple loci in both homozygous and heterozygous conditions. Examination of 15 individual plants within resistant S1 lines, at each of two locations in a previous study (data not presented), suggested that segregation of susceptible plants in highly resistant S1 lines did not occur. In this study, examination of approximately 24 individual plants from each of six S1 lines, and progeny tests in multiple environments, showed no evidence of susceptible segregates. No evidence for a single resistance factor in heterozygous condition was obtained so we concluded that resistance is multigenic. Additionally, we sampled 11 S1 plants of S20 plants and did not detect a non-segregating susceptible S1:2 progeny, providing further evidence (P>/= 0.05) that a homozygous susceptible plant was not present in the S1 progeny (Table 3). Moreover, no chlorotic Ht-type lesions were observed in any progeny. If a dominant, Ht gene were responsible for resistance in these progenies, our sample of six S0 plants should have included at least one heterozygous resistant genotype (P>/= 0.01) (Hanson, 1959). Partially inbred lines can provide family structure for selection in both population improvement and inbred development programmes. Additionally, self-pollination allows deleterious recessive alleles to be revealed and removed from donor populations. Thus, we conclude that S1 lines are useful for both breeding and host resistance objectives. The oligo or polygenically inherited resistance, because of its anticipated durable character, may be very useful for control of NLB. Comparision of resistance responses in different environments showed that high levels of partial resistance alone were sufficient to control the disease in Wooster. Partial resistance genotypes (e.g. Mo17) have been observed to be more susceptible in studies conducted in East Africa (Adipala et al., 1993a; Dingerdissen et al., 1996). The reason for much higher disease development of NLB on Mo17 in East Africa is unclear. Dingerdissen et al. (1996) proposed longer nights or heavier dew (due to lower latitude and higher altitude), plus high humidity from more frequent rains, would better support spore germination and infection in East African environments. Other conceivable reasons include higher amounts of natural inoculum (background) from multiple cropping seasons or higher aggressiveness of East African E. turcicum isolates. ACKNOWLEDGEMENTS We thank Audrey Johnston and Mark Casey for technical assistance, and Lois Zimmerly for preparation of the manuscript. We also thank Kim Campbell for reviewing the manuscript, and Jim Hacker and Burt Bishop for assistance with statistical analyses. Salaries and research support were provided by State and Federal funds appropriated to the Ohio Agricultural Research and Development Center and The Ohio State University, Makerere University, and grant funds from USAID/Uganda Manpower for Agricultural Research and Development (MFAD). REFERENCES Adipala, E., Lipps, P.E. and Madden, L.V. 1993a. Reaction of maize cultivars from Uganda to Exserohilum turcicum. Phytopathology 83:217-223. Adipala, E., Lipps, P.E. and Madden, L.V. 1993b. Occurrence of Exserohilum turcicum on maize in Uganda. Plant Disease 77: 202-205. Campbell, C.L. and Madden, L.V. 1990. Introduction to Plant Disease Epidemiology. John Wiley and Sons, NY. 532 pp. Ceballos, H., Deutsch, J.A. and Gutierrez, H. 1991. Recurrent selection for resistance to Exserohilum turcicum in eight subtropical populations. Crop Science 31:964-971. Dingerdissen, A.L., Geiger, H.H., Lee, M., Schechert, A. and Welz, H.G. Internal mapping of genes for quantitative resistance of maize to Setosphaeria turcica, cause of northern leaf blight, in a tropical environment. Molecular Breeding 2:143-156. Freymark, P.J., Lee, M., Woodman, W.L. and Martinson, C.A. 1993. Quantitative and qualitative trait loci affecting host-plant response to Exserohilum turcicum in maize (Zea mays L.). Theoretical and Applied Genetics 87:537-544. Gevers, H.O. 1975. A new major gene for resistance to Helminthosporium turcicum leaf blight of maize. Plant Disease Reporter 59:296-299. Hanson, W.D. 1959. Minimum family sizes for the planning of genetic experiments. Agronomy Journal 51:711-715. Hooker, A.L. 1961. A new type of resistance in corn to Helminthosporium turcicum. Plant Disease Reporter 45: 780-781. Hooker, A.L. 1967. A second major gene locus in corn for chlorotic-lesion resistance to Helminthosporium turcicum. Crop Science 17:132-135. Hooker, A.L. and Kim, S.K. 1973. Monogenic and multigenic resistance to Helminthosporium turcicum in corn. Plant Disease Reporter 57:586-589. Hughes, G.R. and Hooker, A.L. 1971. Gene action conditioning resistance to northern leaf blight of maize. Crop Science 11:180-184. Jenkins, M.T. 1957. Evaluation of lines for resistance to Helminthosporium. Proceedings Twelfth Annual Hybrid Corn Industry Research Conference. American Seed Trade Association, Chicago, IL. Dec. 4-5, 1957. pp. 7-13. Jenkins, M.T. and Robert, A.L. 1952. Inheritance of resistance to leaf blight of corn caused by Helminthosporium turcicum. Agronomy Journal 44:136-140. Johnson, R. and Jellis, G.L. 1992. Breeding for Disease Resistance. Kluwer Academic Publishers, Dordrecht, The Netherlands. Lipps, P.E., Pratt, R.C. and Hakiza, J.J. 1997. Interaction of Ht and partial resistance to Exserohilum turcicum in maize. Plant Disease 81:277-282. Martin, T.J. and Ellingboe, A.H. 1975. Differences between compatible parasite/host genotypes involving the Pm4 locus of wheat and the corresponding genes in Erisyphe graminis f. sp. tritici. Phytopathology 66:1435-1438. Meyer, A.C., Pataky, J.K. and Juvik, J.A. 1991. Partial resistance to northern leaf blight and Stewart's wilt in sweet corn germplasm. Plant Disease 75:1094-1097. Perkins, J.M. and Pedersen, W.L. 1987. Disease development and yield losses associated with northern leaf blight of corn. Plant Disease 71: 940-943. Pratt, R.C., Ekwamu, A. and Lipps, P.E. 1993. Characterization of race-nonspecific resistance to Exserohilum turcicum races 0 and 1 in maize OhS10 S1 progenies. Plant Disease 77:1227-1232. Pratt, R.C., Findley, W.R., Sotomayor-Rios, A. and Torres-Cardona, S. 1994. Registration of OhS9 (C1) and OhS10 (C1) maize germplasm. Crop Science 34:1418-1419. Raymundo, A.D. and Hooker, A.L. 1981. Measuring the relationship between northern corn leaf blight and yield losses. Plant Disease 65:325-327. Robbins, W.A. and Warren, H.L. 1993. Inheritance of resistance to Exserohilum turcicum in PI 209135, orbelariety of maize. Maydica 38:209-213. SAS Institute, Inc. 1985. SAS Useruide: Statistics. Version 5 ed. SAS Institute, Cary, NC 956 pp. Sigulas, K.M., Hill, R.R. and Ayers, J.E. 1988. Genetic analysis of Exserohilum turcicum lesion expansion on corn. Phytopathology 78:149-153. Steel, R.G.D. and Torrie, J.H. 1980. Principles and Procedures of Statistics. A Biometric Approach. 2nd ed. McGraw-Hill Book Co., NY 633 pp. Turner, M.T. and Johnson, E.L. 1980. Races of Helminthosporium turcicum not controlled by Ht genetic resistance in the American Corn Belt. Plant Disease 64:216-217. Ullstrup, A.J. 1977. Diseases of corn. In: Corn and Corn Improvement. Sprague, G.F. (Ed.), pp. 391-500. American Society of Agronomy, Inc., Madison, WI. Ullstrup, J.M. and Miles, S.R. 1957. The effects of some leaf blights on corn yield. Phytopathology 47:331-336. Welz, H.G., Wagner, R.G. and Geiger, H.H. 1993. Virulence variation in Setosphaeria turcica populations collected from China, Mexico, Uganda, and Zambia. Phytothology 83:1356 (Abstract). Windes, J.M. and Pedersen, W.L. 1991. An isolate of Exserohilum turcicum virulent on maize inbreds with resistance gene HtN. Plant Disease 75:430. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97008c.gif] [cs97008b.gif] [cs97008a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}