 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Short communication Effect of different temperature regimes on vegetative growth of melon plants P. A. AMUYUNZU, J. A. CHWEYA, Y. ROSENGARTNER^1 and S. MENDLINGER^1
Department of Crop Science, University of Nairobi, P. O. Box 29053, Nairobi,
Kenya. (Received 25 March, 1996; accepted 23 November, 1996)
Code Number: CS97010
Sizes of Files:
Text: 25.9K
Graphics: Tables and line drawings (gif) - 42.3K
ABSTRACT Experiments were conducted at the Institutes for Applied Research in Beer-Sheva, Israel between November 1992 and April 1993 to study the effects of different temperature regimes on vegetative growth of thirteen melon (Cucumis melo L.) cultigens and to identify rapid and reliable selection criteria for breeding for cold resistance. The cultigens were grown in greenhouses at two temperature regimes: high and low with mean minimum and maximum temperatures of 13 and 34 C, and 8 and 33 C, respectively. The cultigens were examined for the rate and amount of vegetative growth which included lengths of mainstems, number of nodes, internode lengths and leaf chlorophyll content. Variation among the cultigens both between and within temperature regimes was significant for most of the traits. The best six cultigens for tolerance to suboptimal growing temperatures were Cinco, Midget, BsFDHK, Tal Dvash, H17 and H19. These cultigens were found to perform better, but not necessarily to the same degree in the low temperature regime for all the traits considered than the other cultigens examined. Relative growth rate (RGR) was found to be superior over the rest of the traits in distinguishing between cold sensitive and cold tolerant lines. It could, therefore, serve as a rapid and reliable selection criterion for breeding for cold tolerance in melons. Key Words: Cucumis melo, cold tolerance, cultigen, suboptimal cold temperature RESUME Des experiences sur la croissance de "cultivars" de treize melons (Cucumis melo L.) ont ete menees a l'institut de recherces appliquees de Beer Sheva (Israel) entre novembre 1992 et Avril 1993. Ces experiences ont etudie l'incidence de differents regimes de temperature sur la croissance des cultivars, et cherche a identifier des criteres de selection rapides et fiables pour l'aquisition de la resistance au froid. Les cultigens ont pousse dans des serres a des regimes de temperatures differents; respectivement 1) 13 a 34 C et 2) 8 a 33 C. Ces cultivars ont ete examines en fonction de taux de croisssance vegetative (longueur de la branche centrale, nombre de noeuds, espace entre les noeuds et teneur en chlorophylle). La variation parmi les cultivars entre differents regimes et dans les memes regimes de temperature s'est averee significative pour la plupart des plants. Les six meilleurs cultivars resistants a une temperature suboptimale sont Cinco Midget, BsFDHK, Tal Dvash, H17 et H19. Ces cultivars se sont montres plus performants mais pas necessairement a la meme temperature dans les bas regimes, que les autres plants si l'on distingue la sensation au froid et les lignes de tolerance au froid. Des lors, cela pourrait constituer des criteres de selection rapides et fiables pour acquerir la tolerance au froid. Mots Cles: Cucumis melo, tolerance au froid, cultivars, temperature froide suboptimale, criteres de selection INTRODUCTION Melon (Cucumis melo L.) is an important horticultural crop in many countries of the world. The plants are mainly grown for their edible fruits and, like many other crops of tropical origin, they are adversely affected by exposures to sub-optimal low temperatures (i.e., below 10 C). This is enhanced by the relatively poor cold resistance that exists in the present varieties of the crop. The result of such cold temperature is reduced growth and development of plants which ultimately is followed by extensive reduction in economic yield and quality. The genetic improvement of stress resistance in plants has been a major subject of study because it is an area that is economically important. To date, genetic variation for cold resistance has been identified for various physiological processes but the inheritance of these processes has hardly been studied (Blum, 1988). However, studies by Guy (1990) on the genetics of minimum survivable temperature associated with cold acclimation showed this character to be inherited in a dominant/recessive pattern. In another study it was found that the inheritance of cold germinability in muskmelon (Cucumis melo L.) required the presence of both cytoplasmic factors and nuclear genes and may be under the control of at least four genes (Hutton and Loy, 1992). Therefore, it is possible to improve crops to withstand sup-optimally low temperatures and this is vital in that it will lead not only to yield increase but also to a wider variety of agricultural strategies (Lysons et al., 1979). The objectives of this study were to: (a) investigate the effect of different temperature regimes on vegetative growth of melon plants, and (b) identify rapid and reliable screening procedures for breeding for cold resistance in melons. MATERIALS AND METHODS The field study was conducted at the Institutes for Applied Research in Beer-Sheva, Israel between November 1992 and April 1993. This is an area that is characterised by cold winters between the months of November and March. On average the temperatures during this season ranged from about 5 C at night to 16 C in the day. Temperature regimes. Two temperature regimes for growing melon plants were used. The first was a high temperature regime (T1) provided by heated glass topped greenhouse with two electric heaters whereas the second was a low temperature regime (T2) provided by a greenhouse that was not heated during the winter. The electric heaters in T1 were fitted to a thermostat and set at 25 C. The ambient temperature of the unheated greenhouse approached that of the outside during the night. Temperature in each greenhouse was recorded once a day from two minimum-maximum thermometers, one at ground level and the other placed 1.5 metres high. Plant materials. Seven open pollinated cantaloupe, honey dew, persians and musk-melon types and six F1 hybrid lines produced by crossing the seven were used. The open pollinated cultivars included Haon, BsFDHK, Cinco, Tal Dvash, Crenshaw, Midget and Persian while the F1 hybrids included H2, H13, H14, H17, H18 and H19. The F1 hybrids had been developed in a breeding programme for cold resistance at the Institutes for Applied Research of the Ben Gurion University in Israel. The seven open pollinated lines had previously been identified by the same programme as germplasm that could be useful in improving the cold resistance in melons. Experimental design. The experimental design was a split-plot with temperature regime as the mainplot treatment and type of cultigen as the subplot treatment. There were two temperature regimes, T1 and T2, in heated and unheated greenhouses, respectively, and thirteen cultigens. In each greenhouse, there were nine plants per cultigen. Data collection and analysis. Data were collected on plant height, number of nodes, internode length, length of leaf lamina and leaf chlorophyll content. Height and length of leaf lamina was measured with a transparent plastic ruler. The latter was used in the calculation of Plastochron Index (PI) via Erickson's formulae (Erickson and Michelin, 1957):
n + (ln L(n) - ln (30))
PI = -------------------------
(lnL(n) - ln L(n+1))
where n is the number of leaves on the mainstem which are longer than the reference length of 30 mm, L(n) is the length ogf leaf (n) and L(n+1) is the length of leaf (n+1). PI is used to compare developmental stages of plants at any given time since it is based on the lengths of leaves and their rate of production. (Watts, 1972; Stamp, 1981). Average internode length was calculated on the basis of plant height and number of nodes by dividing the former with the latter. Subsequently, relative growth rate (RGR) was calculated on the basis of mainstem length according to Evans (1972) using the formula:
Ln (L2) - Ln (L1)
RGR = -------------------
T2 - T1
Where L1 is the mainstem length at any one time (T1) and L2 is the mainstem length at time two (T2). Determination of chlorophyll content was based on the work of McKinney (1941) on the absorption of light by aqueous 80% acetone extracts of chlorophyll using a spectrophotometer. Readings for optical density (OD) were made at 663 and 645 nm wavelengths and the total chlorophyll (C) was then calculated using the formula:
(20.2 OD[645] + 8.02 OD[663) x 10]
C = ------------------------------------ mg/g leaf material)
(Leaf weight x 1000)
Relative performance indices. An index (I) was developed to rate the performance of each cultivar in the two temperature regimes using the formula:
c1C2 - c2C1
I = -------------
C1C2
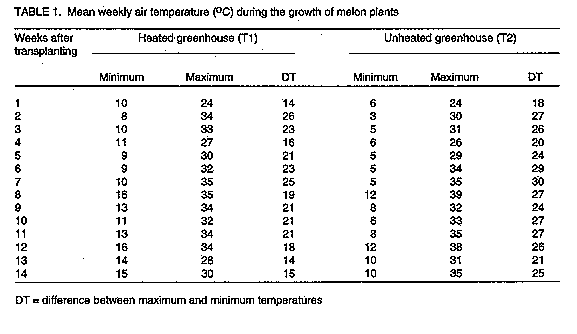
where c1 is the value of a given character for a given cultivar in T1; c2 is the value of the same trait for the same cultivar in T2; C1 is the mean of the trait in T1; and C2 is the mean of the trait in T2 (S. Mendlinger, personal communication). This is an index that was identified (among several of them that had been developed) as being consistent in evaluating the cultivars for cold resistance. Selection of the cultivars for cold resistance was possible using this index because all those cultivars that performed better in the cold temperature regime relative to the high temperature regime assumed negative indices and vice-versa (i.e., cultivars with negative indices performed better in the cold temperature regime while those with positive indices performed better in the high temperature regime). This index compared the performance of each cultivar in the two temperature regimes based on their respective performance in each regime. All data were subjected to appropriate analysis of variance according to Steel and Torrie (1981). RESULTS Temperature. The mean maximum and minimum temperatures for each week of the growing season are listed in Table 1. During the first five weeks, T1, the heated greenhouse temperature was about 2 C warmer in the day and 5 C warmer at night than T2. However, by the start of the 6th week, the day time temperatures between the two greenhouses did not differ but night time temperatures were still lower by 4.5 C in the unheated greenhouse. The high temperature regime, T1, was very similar to that in many commercial greenhouses. Therefore, it would appear that the major differences between the two greenhouses were in the night time temperatures and the total time at suboptimal temperatures.
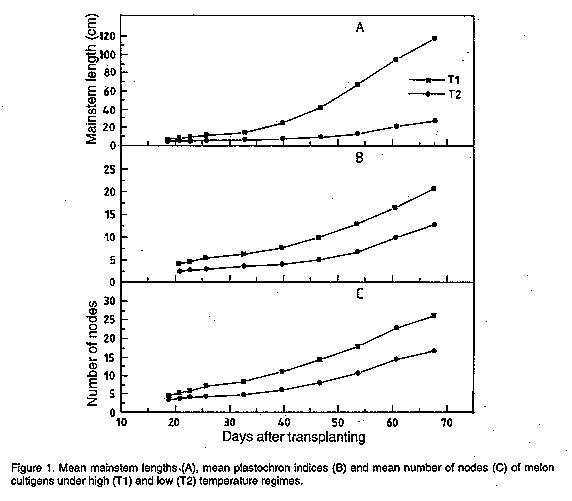
There was also a marked difference between the two temperature regimes in terms of the range between the day's minimum and maximum temperatures, whereby T2 had larger differences than T1. Thus, the plants in T2 experienced greater extremes of temperature than those in T1. Vegetative growth. In general, plants in the two temperature regimes behaved differently vegetatively. There was a general progressive increase in the lengths of the mainstem, number of nodes, internode lengths and PI in both temperature regimes (Fig. 1). The respective values for these vegetative traits at each point in time were generally higher for plants in T1 than those in T2. This difference was most pronounced in the lengths of the mainstems whereby the respective values in T1 were at least thrice those in T2. Subsequently, the rate at which the mainstems increased in length was generally higher in most of the cultivars unlike other traits whose increase was fairly small with time (as depicted in the higher slope of the curve of the former compared to the latter). There was doubling in the lengths of the mainstems of plants growing under T1 weekly.
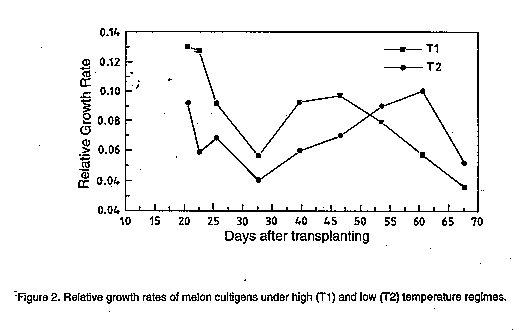
There was no definite pattern followed by RGR of plants in both regimes (Fig. 2). However, plants in T1 had generally higher initial RGR than those in T2. This difference was only evident up to about the seventh week after transplanting at which time the RGR in T1 became lower than that in T2. The combined analysis of variance across the temperature regimes for all the vegetative traits are summarised in Table 2. They showed a significant effect of temperature on all the traits. Also, differences for all traits was significant between cultivars. This suggested the existence of genetic variability among melon cultivars in their capacity to grow at different temperature regimes. There was a substantial genotype-environment interaction effect in all the vegetative traits except in RGR. The various traits had significantly higher values in T1 than in T2, implying a higher vegetative growth in the former than the latter.
Table 3 groups the cultivars into either open pollinated lines or hybrids and compares their performance. For all vegetative traits, except relative growth rate and leaf chlorophyll content, the hybrids had significantly higher values than the open pollinated cultivars. However, significant differences occurred only in the length of the mainstem, number of nodes and internode length. Results of the performance of melon cultivars in the two temperature regimes are presented in Table 4. Of importance are the totals of the various indices of vegetative traits for each cultivar. This gives a measure of performance of each cultivar relative to the two temperature regimes. Therefore, Cinco with a total index of -0.533 performed better vegetatively in T2 while H2 with an index of 0.940 performed poorly. In general, there was no significant difference in the performance of open pollinated lines and hybrids in T2 (Table 5).
DISCUSSION This study clearly shows the effect of temperature on growth and development of melon plants. There was an obvious distinction in the expression of most of the traits whereby plants in the low temperature regime (8 to 33 C) showed marked reduced initial growth and delayed development as a result of the suboptimal temperature stress, in contrast with those in the high temperature regime (13 to 34 C). This difference in growth and developmental patterns associated with temperature emphasises the important role played by temperature in all the physiological and biochemical processes in plants. Therefore, the ability of a crop plant to minimise the unfavourable effects induced by cool temperature is an important attribute advantageous to plant survival, maturition and yield (Pozzi et al., 1986). Initial vegetative growth of the plants was highly influenced by genotype and temperature regimes. There were distinct differences between the genotypes in the parameter response to temperature. This implied that all vegetative traits are genetically controlled, and that melon genotypes differ in this respect. In general, high temperature regime led to a vigorous vegetative growth of the plants in contrast to the low temperature regime. Plants under the higher temperature regime had generally longer mainstems and internode lengths, and many nodes compared to those under the lower temperature regime. This is consistent with results obtained by Slack and Hand (1983) in cucumbers, Malek et al. (1992) in flame azelea, Grimstad (1993), and Grimstad and Frimanslund (1993) in cucumbers. The difference in vegetative behaviour between the two temperature regimes is reflected in the difference in relative growth rates in which plants in the higher temperature regime had a considerably higher initial relative growth rate than those in the lower temperature regime. This disparity in RGR was probably due to the difference in the maturity of plants whereby those in T1 grew faster and, hence, attained maturity earlier than those in T2. Similarly, the difference in rates of plant growth was indicated by the Plastochron Index in which case a high index indicated an advanced stage of development (Silk, 1980). Thus, a plant which has five leaves after one week at 25 C, will be morphologically and biochemically identical to a plant which has five leaves after two weeks at 15 C. As such, at any time plants in the higher temperature regime were physiologically older than those in the lower temperature regime. Another important character reflected in Plastochron Index is the vegetative framework, especially the number and area of leaves. A large PI implies a greater number and area of leaves and, therefore, plants in the higher temperature regime had generally higher number of leaves compared to those in the lower regime. This implies that plants in T1 had a better vegetative establishment than those in T2. A lower leaf number and area in the cold greenhouse may have been due to cold temperature stress (Blum, 1988; Aoki et al., 1989). A poor vegetative establishment of plants in T2 implied a limited available photosynthetic surface and, hence, the rate and amount of assimilate production may have been poor in this regime. In contrast, plants in T1 had a greater surface area for photosynthesis and this may have enhanced the rate of assimilate production. Generally, a high rate of assimilate production implies a high rate of plant growth and, therefore, the observations that were made in both temperature regimes concided with this. These observations support those of Friend and Helson report (1976) in which high temperature regime was found to favour a high rate of photosynthesis in plants. Subsequently, assimilate production in the higher temperature regime may have been enhanced by the relatively high amounts of total chlorophyll per unit leaf weight, which are important pigments during the process of photosynthesis. Therefore, the cold temperature stress to which the plants were subjected to in T2 resulted in lower amounts of chlorophyll per unit leaf weight. This is consistent with studies on cold stress done by Mellerd and McWilliam (1968) in maize, Kabaki et al. (1983) in rice, and Aoki et al. (1989) in cucumber. These studies showed that plants grown under cold temperature regimes had significantly lower rates of photosynthesis as a result of low amounts of leaf chlorophyll. There was marked variation in the resistance of the various genotypes to cold in terms of vegetative growth. The genotypes which were superior in the cold regime and, hence, showed increased tolerance to cold included Cinco, Midget, BsFDHK, Tal Dvash, H17 and H19. The other genotypes performed poorly in this regime. On average, there was no significant difference in performance between open-pollinated and hybrid lines in the low temperature regime. However, in terms of the general plant anatomy, hybrids were generally larger in size than the open pollinated plants. For the purpose of this study, these results showed that most of the vegetative variation that was observed in the melon plants was due to the influence of genotype and temperature. However, while most of the traits showed genotype-environment interaction, RGR did not. This implies that the differences that were observed in RGR between the two regimes was mainly due to the difference in the ambient temperature. Therefore, this trait appears to be an indicator of plant tolerance to periods of unfavourable cool conditions. However, the use of vegetative traits in general as screening criteria may be trivial, especially as far as fruit biomass production is concerned. This is so because, in an intensive agricultural system, the source (leaf area) is usually large compared with its potential sink (i.e., the fruit) and, therefore, a reduction in vegetative growth may be unimportant for fruit biomass production (Mendlinger and Pasternak, 1992). ACKNOWLEDGMENTS The authors thank Mr. Haim Koperman of the Institutes for Applied Research, Ben-Gurion University of the Negev for his moral and technical support. The study was sponsored by the Division for International Cooperation (Mashav) of the Ministry of Foreign affairs of the State of Israel. REFERENCES Aoki, S., Oda, M. and Hosino, K. 1989. Varietal differences in chilling-induced depression of photosynthesis and leaf growth in cucumber seedlings. Journal of Japanese Society of Horticultural Science 58:173-179. Blum, A. 1988. Plant Breeding for Stress Environments. CRC Press Inc, Boca Raton. Florida. 199-110. Erickson, R. O. and Michelini, F. J. 1957. The plastochron index. American Journal of Botany 44:297-305. Evans, G. C. 1972. Studies in ecology. Vol. 1. The Quantitative Analysis of Plant Growth. Blackwell Scientific Publicattions, Osney Mead, Oxford, England. Friend, D. J. C. and Helson, V. A. 1976. Thermoperiodic effects on the growth and photosynthesis of wheat and other crop plants. Botanical Gazette 137:75-84. Grimstad, S. O. and Frimanslund, E. 1993. Effect of different day and night temperature regimes on greenhouse cucumber young plant production, flower bud formation and early yield. Scientia Horticulturae 53:191-204. Grimstad, S. O. 1993. The effect of a daily low temperature pulse on growth and development of greenhouse cucumber and tomato plants during propagation. Scientia Horticulturae 53:53-62. Guy, C. L. 1990. Cold acclimation and freezing stress tolerance: role of protein metabolism. Annual Review of Plant Physiology Molecular Biology 41:187-223. Hutton, M. G. and Loy, J. B. 1992. Inheritance of cold germinability in muskmelon. HortScience 27:826-829. Kabaki,N., Tajima, K. and Amemiya, A. 1983. Physiological study of growth inhibition in rice as affected by low temperature. 1. Physiological mechanisms of growth retardation caused by cool temperature. Bulletin of the National Institute of Agricultural Sciences of Japan. D, 34, 1. Lyons, J. M., Graham, D. and Raison, J. K. (Eds.). 1979. Low Temperature Stress in Crop Plants: The Role of the Membrane. Academic Press, New York. 565pp. MacKinney, G. 1941. Absorption of light by chlorophyll solutions. Journal of Biochemistry 140:315-322. Malek, A. A., Blazich, F. A., Warren, S. L. and Shelton, J. E. 1992. Initial growth of seedlings of flame azelea in response to day/night temperature. Journal of American Society of Horticultural Science 117:216-219. Millerd, A. and McWilliam, J. R. 1968. Studies on a maize mutant sensitive to low temperature. Plant Physiology 43:1967-1972. Mendlinger, S. and Pasternak, D. 1992. Effect of time of salinization on flowering, yield and fruit quality factors in melon, Cucumis melo L. Journal of Horticultural Science 67:529-534. Pozzi, G. L., Gentinetta, E., Salamini, F. and Motto, M. 1986. Genotypic variation for cold tolerance among twelve maize (Zea mays L.) populations. Plant Breeding 97:2-12. Silk, W. K. 1980. Plastochron indices in cantaloupe grown on an irrigation line source. Botanical Gazette 141:73-78. Slack, G. and Hand, D. W. 1983. The effect of day and night temperatures on the growth, development and yield of glasshouse cucumbers. Journal of Horticultural Science 58:567-573. Stamp, P. 1980. Activity of photosynthetic enzymes in leaves of maize (Zea mays L.) in relation to genotype and to temperature changes. Z. Pflanzenphysiol. 100:229-239. Steel, R. G. D. and Torrie, J. H., 1981. Principles and Procedures of Statistics. A Biometrical Approach. McGraw-Hill Co. 377-398. Watts, W. R. 1972. Leaf extension in Zea mays L. II. Leaf extension in response to independent variations of the temperature of the apical meristem, of the air around the levels, and of the root zone. Journal of Experimental Botany 23:713-721. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97010c.gif] [cs97010g.gif] [cs97010f.gif] [cs97010d.gif] [cs97010b.gif] [cs97010a.gif] [cs97010e.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}