 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 5. No. 2, pp. 135-146, 1997 Grain yield of barley as affected by cropping sequence and fertilizer application in southeastern Ethiopia ASEFA TAA, D.G. TANNER^1, KEFYALEW GIRMA and AMANUEL GORFU
Kulumsa Research Centre, P.O. Box 489, Asella, Ethiopia (Received 9 November, 1996; accepted 18 March, 1997)
Code Number: CS97020
Sizes of Files:
Text: 41.6K
Graphics: Line drawings (gif) - 20.6K
ABSTRACT Cropping systems in the Ethiopian highlands involve rotations of cereals with grain legume and oilseed crops; the proportional allocation among crop species varies with altitude, rainfall, and soil type. Barley (Hordeum vulgare) tends to dominate in the highest altitudinal zones, while bread wheat (Triticum aestivum) is more common at medium altitudes. A trial was established in 1992 at the Bekoji research site in southeastern Ethiopia to evaluate interactions among barley-based cropping sequences and annual applications of inorganic nitrogen (N) and phosphorus (P) fertiliser. Rotational crops included Ethiopian rapeseed (Brassica carinata), faba bean (Vicia faba), and bread wheat. Results indicated that barley grain yield (GY) was significantly affected by crop rotation, fertiliser P, and by rotation by P interaction. Barley GY in rotations with dicotyledonous crops exceeded that in cereal rotations by 62 and 46% in the second and third seasons. Barley GY in two year rotational sequences exceeded three year sequences by 31%; within the three year sequences, the GY of the first barley crop exceeded that of the second crop by 57 %. There was a significant decline in barley GY upon extending the dicot rotations to three years from two (-868 kg ha^-1), while extending the cereal rotation had no effect on barley GY. In 1994, rotation by P interaction revealed a greater P response in dicot vs. cereal rotations: in the former, GY response to 20 kg P was 29.8 kg grain per kg P, while in the cereal rotation the ratio was only 14.4:1. Grass weed densities were affected by rotation cycle duration and by P application. Density of grass weed inflorescences, mostly Avena fatua panicles, at maturity (mean 29 m^-2) was significantly higher in second crop barley in three year rotations, and was lowest in first crop barley without N fertiliser. Grass inflorescence density was significantly lower in P fertilised plots. Key Words: Barley, crop rotation, faba bean, grain yield, nitrogen, phosphorus, rapeseed, weed density, wheat RESUME Les systemes de culture sur les hauts plateaux ethiopiens reposent sur une rotation cereales-legumineuses-oleagineux. Les proportions relatives des differentes especes varient en fonction de l'altitude, de la pluviometrie, et la qualite du sol. La culture d'orge (Hordeum vulgare ) domine dans les parties les plus hautes, tandis que le ble (Triticum aestivum) se rencontre plutot aux altitudes intermediaires. Un dispositif experimental a ete mis en place en 1992 au centre de recherche de Bekoji (dans le sud-est ethiopien), afin d'evaluer, l'interaction entre le schema de rotation et l'application annuelle d'engrais inorganique, azote (N) et phosphore (P) au sein des systemes a base d'orge. Le colza ethiopien (Brassica carinata), la feve (Vicia faba), et le ble tendre ont ete utilises comme cultures de rotation. Les resultats montrent un effet significatif du type de rotation, de la fertilisation P, ainsi que de l'interaction rotation-fertilisation P sur le rendement en grains de l'orge (RG). Le RG de l'orge en rotation avec des dicotyledones est superieur a celui de l'orge en rotation avec des cereales. La difference est de 62 % et 46% pour les deuxieme et troisieme cycles de culture respectivement. Le RG de l'orge en rotation sur deux ans est superieur de 31% a celui de l'orge en rotation sur trois ans. Si l'on considere les rotations sur trois ans, le RG de l'orge de premiere annee est superieur de 57% a celui de l'orge de deuxieme annee. On observe une reduction significative du RG si l'on prolonge la rotation avec des dicotyledones de deux a trois ans (-868 kg ha^-1). En revanche, l'extension de la rotation avec des cereales n'a pas d'effet sur le RG. Des essais d 'interaction rotation-fertilisation P menes en 1994 montrent que la reponse a la fertilisation Pest plus importante dans le cas de rotations avec des dicotyledones que dans le cas de ceiles avec des cereales: dans le premier cas, la reponse du RG a 20 kg de Pest de 29.8 kg de grains par kg de P, tandis que dans le deuxieme, le ratio n'est que de 14.4 pour 1. La duree de la rotation et de la fertilisation Pont par ailleurs un impact sur la densite de graminees adventices. La densite d'inflorescences d'adventices a maturite est en moyenne de 29 par metre carre (il s'agit essentiellement de panicules d'Avena fatua). Elle est significativement plus elevee en deuxieme annee d'orge dans le cas d'une rotation sur trois ans; le niveau minimum correspond au cas d'un orge de premiere annee sans fertilisation azotee. La densite de graminees adventices est significativement plus faible dans les parcelles ayant subi une fertilisation P. Mots Cles: Azote, ble, colza, densite d'adventices, feve, orge, phosphore, rendement en grains, rotation culturale INTRODUCTION The maintenance of long-term agricultural productivity in any ecosystem depends on a number of biotic and abiotic factors all of which are dynamic in response to human intervention. Conservation tillage and crop rotation are considered to be major means of maintaining agricultural productivity globally (Lal, 1989). Historically, few agricultural research programmes in sub-Saharan Africa (SSA) have conducted multi-year or long-term agronomic trials to investigate the impact of crop production practices on the sustainability of the prevailing farming systems.
Practical long-term rotation options for cropping systems in barley producing zones have seldom been reported in SSA. However, one study in Ethiopia considered the benefits from crop rotation, examining the impact of various cropping sequences on barley grain yield (Amanuel et al., 1994).
Fertiliser rates have often been recommended for crop production in SSA without considering the sustainability of continuous fertiliser application. In Ethiopia, the high rates of nitrogen (N) applied to bread wheat plots in single season on-farm fertiliser trials had repercussions on soil pH levels, severity of wheat foliar disease, and weed incidence and competition (Tanner et al., 1993). However, there are no reports available on the long-term effects of repeated N and phosphorus (P) fertiliser application at the rates recommended for barley (Hordeum vulgare L.) production.
In the peasant farming systems of southeastern Ethiopia, cereals predominate, often occupying over 80% of the total cropped land each season (Chilot et al., 1992). In the highland zones, bread wheat (Triticum aestivum) and barley are the most common cereals in production, while faba bean (Vicia faba) and rapeseed (Brassica carinata) are common grain legume and oilseed crops, respectively. The mean area under barley production in Ethiopia during the period 1980-89 has been estimated at 892,500 ha (Berhane et al., 1996).
The high proportion of wheat and barley in the highland cropping systems satisfies the shortterm subsistence objectives of peasant farmers, but may prove disadvantageous in the long-term due to the absence of the inherent advantages of crop rotational systems. Several of the well-known benefits of crop rotation could be extremely valuable in the peasant farming sector: N fixation by legumes (Hargrove et al., 1983); the interruption of weed (Heenan et al., 1990), disease (Cook, 1984) and insect cycles by dicotyledonous crops; crop diversification (Zentner and Campbell, 1988); improvement in soil tilth and a concomitant reduction in rainfall runoff and erosion (Higgs et al., 1990).
Several short-term studies in the Ethiopian highlands have examined the beneficial effects of break crops on bread wheat production. In one study, a faba bean break crop increased wheat grain yield by 1100 kg ha^-1, or 69% of the yield of second year continuous wheat (Hailu et al., 1989); in a second study, faba bean increased wheat yield by 1000 kg ha^-1, or 44% of. the yield of continuous wheat (Asefa et al., 1992). However, no studies have previously considered the effects of diverse cropping sequences on the performance of barley in the Ethiopian cropping environment.
This paper presents the results generated during 1992-94 of one such trial in a major barley production zone in southeastern Ethiopia. The objective of this trial was to evaluate the effects of alternate crop rotation systems on barley productivity and on barley response to inorganic fertiliser. MATERIALS AND METHODS The rotation experiment was established in 1992 at the Bekoji (7 degrees 34'N and 39 degrees 15'E) research site in Arsi Region of Ethiopia. The station soil is classified as an eutric Nitosol, exhibiting a clay content of approximately 48.5% (Tanner et al., 1993), and is relatively deficient in P. The characteristics of the 0-20 cm soil layer include a pH of 5.1-5.5, a P (Mehlich) content of 7.5-8.2 ppm, and an organic matter content of 4.7-6.3% (Amanuel et al., 1994). The average monthly mean minimum and maximum temperatures during the annual growing season are 7.4 and 17.9 C, respectively. Total annual precipitation is 1048 mm, and that received during the June-November growing season is 657 mm (mean of 1972-95). The station is situated at an altitude of 2780 m a.s.l. The experimental site used for the rotation trial had been sown to an unfertilised barley crop in 1991.
The rotation trial incorporates three fundamental design principles consistently stressed in the statistical literature on multi-rotation experiments (Patterson, 1965; Preece, 1986; Cady, 1991):
Each phase of a specific rotation is present in the experiment every year to avoid assessing the effects of the rotational crops under differing seasonal conditions (i.e., confounding treatment and year effects). A three year cycle requires three plots in each replication to include all three phases of the rotation, while a two year cycle occurs in two phases each year.
Randomisation is essential to avoid anomalies from systematic assignment of treatments to field plots, and is fundamental to the validity of statistical analyses.
Replication is required to estimate experimental error and to control the magnitude of the standard error of treatment means.
The experiment was laid out in a split-plot design with crop rotations as main plots and fertiliser levels as subplots. The crop rotations are barley-based, and reflect the predominant crops in the surrounding farming system: faba bean, rapeseed and bread wheat. Treatments 1 to 15 (Table 1) comprise all phases of barley in two and three year rotations with the three break crops, while treatment 20 consists of continuous barley. Treatment pairs 16 and 17 and 18 and 19 comprise a partial sampling of all potential phases of complex six and four year rotations, respectively. The four fertiliser levels used are the factorial combinations of O and 41 kg N ha^-1 with 0 and 20 kg P ha^-1. The standard recommended rate of macronutrient application for barley production in Ethiopia is 41 kg N and 20 kg P ha^-1 (Taye et al., 1996). The four fertiliser treatments are being maintained as fixed subplots over the duration of the trial. Urea and triple superphosphate are the sources of N and P, respectively. The trial was laid out in three replications, and the area of each main plot is 100 m^2 with each fertiliser subplot having an area of 25 m^2.
TABLE 1. Treatments included in the barley-based crop rotation experiment at Bekoji ------------------------------------------- Treatment Rot phase 1992 1993 1994 ------------------------------------------- 1 1-1 Fb* Ba Fb 2 1-2 Ba Fb Ba 3 2-1 W Ba W 4 2-2 Ba W Ba 5 3-1 R Ba R 6 3-2 Ba R Ba 7 4-1 R Ba Ba 8 4-2 Ba Ba R 9 4-3 Ba R Ba 10 5-1 Fb Ba Ba 11 5-2 Ba Ba Fb 12 5-3 Ba Fb Ba 13 6-1 W Ba Ba 14 6-2 Ba Ba W 15 6-3 Ba W Ba 16 7-1 Fb Ba Ba 17 7-2 R Ba Ba 18 8-1 Fb Ba R 19 8-2 R Ba Fb 20 9 Ba Ba Ba * Fb, W, Ba, R = faba bean, bread wheat, barley, and rapeseed, respectively --------------------------------------------Tillage for this trial is based on the local ox-plough; crop protection practices simulate farmer practices (i.e., hand weeding); varietal selection is optimal for each crop, and is open to change over the trial duration. The recommended varieties used for the various crop species were: for bread wheat, Enkoy in 1992-93 and HAR 1709 in 1994; for barley, Holker in 1992-93 and ARDU 12-60B in 1994; for faba bean, CS20DK in 1992-94; for rapeseed, Yellow Dodola in 1992-94. The seed rates used were 150 kg ha^-1 for bread wheat, 200 kg ha^-1 for faba bean, 15 kg ha^-1 for rapeseed, and 85 (in 1992 and 1993) and 130(in 1994) kg ha^-1 for barley. Pre-weighed amounts of seed and fertiliser were broadcast-applied on the soil surface of the appropriate plots after tillage by ox-plough, and were subsequently incorporated, in the traditional Ethiopian practice, by one pass of the ox-plough. Sowing dates were June 25, 1992, June 23, 1993, and June 21, 1994.
Data were collected from each subplot on days to heading and maturity, plant height, and weed density. At maturity, a net plot of 9 m^2 was harvested by sickle at ground level for grain and biomass yield determination. Grain moisture contents were determined and yields adjusted to a 12.5% moisture basis for wheat and barley, 7% for rapeseed, and 10% for faba bean.
All data were subjected to ANOVA, analysing the four crop species separately. Subsequently, for the barley data of 1993 and 1994, orthogonal contrasts were used to partition rotation main effects and rotation by fertiliser interaction effects into meaningful single degree of freedom components. In 1994, rotation (with 11 degrees of freedom) was partitioned into the following eight mutually orthogonal contrasts:
2) barley in 2 year vs. 3 year rotation; 3) within 3 year rotations, first crop barley vs. second crop barley; 4) barley in rotation with faba bean vs. rapeseed; 5) interaction of contrasts 1 and 2; 6) interaction of contrasts 2 and 4; 7) interaction of contrasts land 3; 8) interaction of contrasts 3 and 4.
Rotation by fertiliser interaction (with 33 degrees of freedom) was partitioned into eight mutually orthogonal contrasts:
3-4) (barley in 2 year vs. 3 year rotation) x (N or P effect); 5-6) (within 3 year rotations, first crop barley vs. second crop barley) x (N or P effect) 7-8) (barley in rotation with faba bean rs. rapeseed) x (N or P effect).
Seed yields of the four crop species included during the first three cycles of the crop rotation trial are summarised in Table 2, including N and P effects on the seed yield (SY) of all species, and rotation main and interaction effects for barley. TABLE 2. Yields (kg ha^-1) of four crops included in the barley-based crop rotation trial at Bekoji and significance levels of treatment effects
-------------------------------------------------------
1992 1993 1994
-------------------------------------------------------
Faba bean Mean yield 2306 3133 2075
C.V.(%) 25.5 15.1 26.7
N effect * NS NS
P effect ** * **
Rapeseed Mean yield 2186 2036 2960
C.V.(%) 34.5 23.2 12.2
N effect NS NS NS
P effect *** - -
Wheat Mean yield 3054 1399 75 5
C.V.(%) 20.5 41.4 34.1
N effect NS NS NS
P effect NS NS NS
Barley Mean yield 1661 1691 1063
C.V.(%) 19.7 15.5 36.3
Rotation effect - *** **
N effect NS ** NS
P effect *** *** ***
Rot. x N interaction - NS NS
Rot. x P interaction - * NS
*, **, ***: Significant at the 5, 1 and 0.1% level, respectively
NS: Not significant (P>0.1)
--------------------------------------------------------
Faba bean (Fb) responded marginally to applied N in 1992 (i.e., 8.7 kg seed per kg N applied), but not at all in 1993-94. Faba bean exhibited a marked response to P application in all three cycles: P use efficiency ratios were 28.1,43.3 and 39.7 kg of seed per kg of fertiliser P across the three seasons. On the plots which did not receive P for three consecutive seasons, the SY of Fb was 1679 kg ha-1 or 68% of the Fb yield on plots receiving 20 kg P ha^-1 annually.
A rotation effect on Fb SY was apparent during 1994, the only season in which Fb was grown subsequent to different precursor sequences. Faba bean grown after Fb 2 years previous (i.e., with an intervening barley crop) yielded significantly lower (1528 kg ha^-1) than Fb after rapeseed 2 years previous (2407 kg ha^-1) or Fb after 2 consecutive crops of barley (2291 kg ha^-1). This yield depression reflects a potentially negative aspect of including Fb too frequently in a barley-based rotation (i.e., 1 year in 2), and may suggest a negative effect on crop root health (Cook, 1984), resulting from over-exploitation of a particular soil zone and/or a build-up of specific root pathogens.
Rapeseed responded positively to fertiliser P in 1992 (Table 2), exhibiting a P conversion efficiency of 15.6 kg seed per kg P applied. In 1992, rapeseed on the zero P plots produced a mean SY of 2031 kg ha^-1. However, in 1993 and 1994, rapeseed sown on the zero P subplots (i.e., subplots receiving zero P in each year of the trial) exhibited poor emergence, reduced plant vigour and stunted growth throughout its development, and produced essentially no SY in both seasons. During the same two seasons, rapeseed sown on plots receiving an annual application of 20 kg P ha^-1 produced SYs of 2036 and 2960 kg ha^-1. As a result, the zero P rapeseed subplots were omitted from the data analysis in 1993 and 1994, but N response was calculated each season by contrasting the yield of rapeseed on the subplots receiving 41-20 and 0-20 kg N-P ha^-1 annually. Thus, of the four crop species included in this trial, rapeseed proved to be the most sensitive to P application.
In contrast, bread wheat did not exhibit a response to N or P in any of the three seasons (Table 2). Mean wheat grain yield (GY) was markedly reduced (and the trial C.V. increased) in 1993 by the outbreak of a severe epidemic of a new race of stem rust (Puccinia graminis f. sp. tritici) to which the variety Enkoy was highly susceptible. As a result, another wheat variety, HAR 1709. was substituted in the trial in 1994, but this variety exhibited poor adaptation and a low GY at Bekoji. Thus, the absence of either N or P response in 1993-94 in bread wheat must be interpreted in the context of poorly-adapted genotypes.
Barley, the main component of the trial, performed relatively consistently across the first three seasons (Table 2). Barley GY response to 41 kg N ha^-1 was significant only in 1993, but with a low conversion efficiency of 3.1 kg grain per kg N applied. Consistent with the results obtained in Fb and rapeseed, barley GY exhibited a significant response to the annual application of 20 kg P ha^-1 in all three seasons; P conversion efficiency was 30.9, 13.7 and 24.7 kg grain per kg fertiliser P during 1992-94, respectively. In 1994, barley GY on the plots receiving 0 P for three consecutive seasons was 817 kg ha^-1 or 62% of the yield level of treatments receiving 20 kg P ha^-1 annually.
Thus, the magnitude of the response to P on this relatively infertile soil was consistent with a previous report (Amanuel et al., 1994) in which P response was the overriding factor affecting crop yields at Bekoji. Rotation effects on barley GY. In 1993, the rotation main effect on barley GY was highly significant (Table 2). Upon partitioning of the rotation main effect, three components were found to be significant:
2) GY of barley following wheat (1595 kg ha^-1) was 48% higher (P<0.001) than the GY of a second consecutive barley crop (1078 kg ha^-1); and 3) the GY of barley after faba bean (1925 kg ha^.t) was 9% lower (P) than after rapeseed (2115 kg ha^-1).
The current results differ from those of a previously reported trial at Bekoji (Amanuel et al, 1994) in which cropping sequence had only a marginal impact on barley GY. Furthermore, in the previous trial, barley following faba bean tended to out-perform barley after rapeseed. Research reported elsewhere indicates a predominantly positive influence of leguminous precursor crops on subsequent cereal crops (Hargrove et al., 1983; Narwal et al., 1983; Hailu et al., 1989; Higgs et al., 1990; Franzluebbers et al., 1995).
In 1994, the effect of rotation on barley GY was also highly significant (Table 2). Partitioning of the rotation main effect revealed five significant components:
2) barley GY in 2 year (cycle) rotations ( 1447 kg ha^-1) was 31% higher (P<0.05) than in 3 year rotations (1105 kg ha^-1); 3) GY of the first barley crop after any precursor crop (1350 kg ha^-1) was 57% higher (P<0.05) than the GY of the second consecutive barley crop (861 kg ha^-1); 4) the interaction of dicot vs. cereal rotation by first rs. second barley crop affected (P<0:O01) barley GY; and 5) the interaction of dicot vs. cereal rotation by 2 vs. 3 year rotations affected (P<0.1) barley GY.
In 1994. there was no significant difference between the effects of faba bean and rapeseed precursor crops on barley GY; this contrasts with the 1993 results, but agrees with the previous published report (Amanuel et al., 1994).
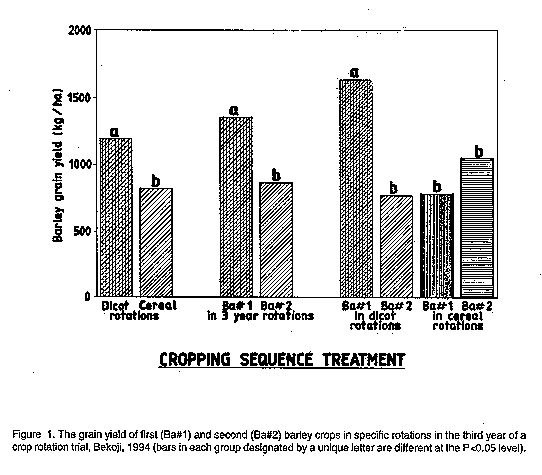
The rotation interaction effects on barley GY are shown in Table 3. The interaction between rotational crop (i.e., dicot vs. cereal) and rotational sequence (i.e., first or second barley crop after the precursor) arose from the following (Table 3): the beneficial effect of the dicot rotation on barley GY was only observed in the first [Ba#1] subsequent barley crop; the GY of the second [Ba#2] consecutive barley crop after either dicot precursor was not different from either Ba#l or Ba#2 after a cereal precursor. Thus, extending the cycle length of a barley rotation with dicots to 3 years (i.e., 2 barley crops + 1 dicot crop) from 2 years resulted in a significant reduction in barley GY of 868 kg ha^-1 in the second barley crop.
TABLE 3. Effects of interaction of rotational crop by sequence on barley grain yield (kg ha^-1) at Bekoji, 1994
--------------------------------------------------------------
Within 3 y cycle Cycle length (y)
Figure 1 illustrates the relation between the rotation interaction and main
effects on barley GY: the superiority of dicot vs. cereal rotations and
Ba#l vs. Ba#2 in 3 year rotations is primarily due to the enhanced
performance of Ba#1 within dicot rotations.
The interaction (P<0.1) between cycle length and rotational sequence (Table 3) is evident in the ranking of mean barley GY for the four combinations: 2 year dicot rotations > 3 year dicot rotations > 2 or 3 year cereal rotations.
Cropping sequence effects on barley GY in 1993 and 1994 are summarised in Table 4. The indexed values highlight the superior GY performance of barley sown immediately subsequent to either dicot crop - an advantage which largely dissipates in the second consecutive barley crop. In contrast to the previous report (Amanuel et al., 1994), barley GY is enhanced, albeit to a lesser extent, by rotation with wheat; this may be due to a disruption of the life cycles of barley root pathogens (Cook, 1984), or could even reflect the residual effect of the low-yielding bread wheat variety, resulting in low nutrient extraction, in 1993. TABLE 4. Barley grain yield response to break crops as a percent of the yield of continuous barley at Bekoji, 1993 and 1994
------------------------------------------
1993 1994
------------------------------------------
First year after faba bean 179 303
Second year after faba bean - 118
First year after rapeseed 197 299
Second year after rapeseed - 128
First year after wheat 148 147
Second year after wheat - 188
Continuous barley 100a 100b
a, b Index yields = 1078 and 560 kg ha-1, respectively
------------------------------------------------------
Rotation by nutrient interaction. Although rotation by N interaction was
not significant in both 1993 and 1994, and rotation by P interaction was
also not significant in 1994 (Table 2), four individual orthogonal
contrasts were significant over the 2 years:
2) (faba bean vs. rapeseed precursor) by N (P<0.1 ) in 1993; 3) (dicot vs. cereal rotation) by P (P<0.001) in 1993; and 4) (dicot vs. cereal rotation) by P (P) in 1994.
The significant interaction between (dicot vs. cereal rotation) by N application (Table 5) indicated that the GY of barley in rotation with dicots showed no response to an annual application of 41 kg N ha^-1; in cereal rotations, barley GY exhibited a significant response to N. but with a conversion efficiency of only 5.8 kg grain per kg N applied. Barley GY responded to the combined effects as follows: dicot+N = dicot-N > cereal+N > cereal-N. The magnitude of the rotation effect on barley GY exceeded that of applied N fertiliser. The contribution of additional N to the cereal by leguminous precursor crops has been termed the "N effect" while yield benefits beyond that accounted for by N was referred to as "rotation effect" by Franzluebbers et al. (1995). Clearly, as shown by the 1993 results, the "rotation effect" on barley GY was more important than the"N effect" at Bekoji.
The marginally significant (P<0.1 ) interaction of faba bean vs. rapeseed precursor crop by N application is somewhat puzzling (Table 5). For barley following rapeseed, there was no GY response to applied N, but, following faba bean, there was a significant GY response to N, but with a low conversion efficiency of only 3.3 kg grain per kg N applied. The apparent lack of a "N effect" of the faba bean precursor. a highly productive leguminous crop (Table 2), disagrees with the published literature stating that cereal GY response to fertiliser N and optimal application rates are normally reduced following grain legume precursor crops (Hargrove et al., 1983; Narwal et al., 1983; Ladd and Amato, 1985; Hailu et al., 1989; Franzluebbers et al., 1995). This apparent contradiction is currently the subject of a more intense investigation at Bekoji, using rhizobial inoculants and N^15 labelling. TABLE 5. Effects of interaction of fertiliser N application by rotational crop on barley grain yield (kg ha^-1) at Bekoji, 1993
----------------------------------------------------
Rotational crop Dicot Cereal Faba bean Rapeseed
----------------------------------------------------
N fertiliser:
- 2000 1132 1856 2145
+a 2044 1369 1993 2095
LSD (P=0.05) 123 126b
a 41 kg N ha^-1
b LSD at P=0.10 level
----------------------------------------------------
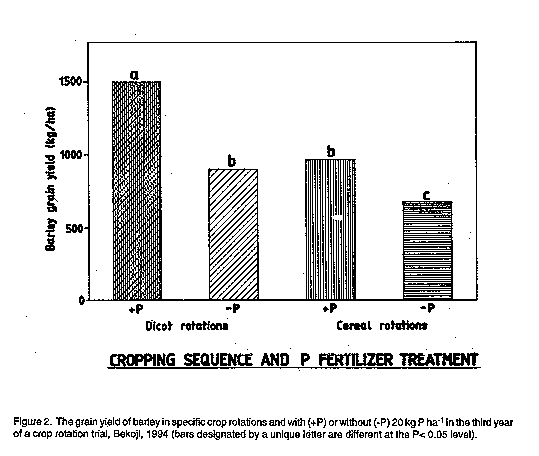
The rotation by P interaction effects on barley GY in both 1993 and 1994
(Table 6) were similar in that, in both seasons, the dicot + P treatment
outperformed the other 3 combinations, and the cereal - P treatment was
significantly lower than the other 3 combinations. In 1993, the conversion
efficiency was higher for barley in cereal rotations (22.3 kg grain per kg
P applied) vs. in dicot rotations (7.2 kg grain per kg P). In 1994, this
order was reversed with a higher conversion efficiency exhibited by barley
in dicot rotations (29.8 kg grain per kg P) vs. in cereal rotations 14.4 kg
grain per kg P). These contrasting results are due to differences in
treatment ranking: in 1993, dicot - P > cereal + P, whereas in 1994. dicot
P = cereal + P. Thus, the pronounced beneficial impact of dicot rotations
on barley GY in 1993 apparently more than compensated for the absence of P
fertiliser. The 1994 results (Fig. 2) are more representative of other
reports in the literature (Sinha et al., 1983) of enhanced P
use-efficiency in legume-cereal rotations.
----------------------------------------------
Rotational crop 1993 1994
----------------------------------------------
Dicot Cereal Dicot Cereal
-------------- --------------
P fertiliser:
- 1951 1028 899 671
+a 2094 1473 1485 958
LSD (P=0.05) 123 222
a 20 kg P ha^-1
----------------------------------------------
N, P and rotation by P interaction were nonsignificant. However, rotation
by N interaction significantly (P<0.05) affected broadleaf seedling
density. The highest density of broadleaf weed seedlings occurred on Dicot
+ N plots, being significantly higher than Dicot - N or Cereal + N (Table
7).TABLE 7. Effects of interaction of fertiliser application by rotational sequence on weed density in barley at Bekoji, 1994
---------------------------------------------------------------------------
Rotational sequence Total broadleaf seedling Total grass inflorescence
density^b density^c
------------------------- -------------------------
Dicot Cereal Ba#1^d Ba#2
---------------------------------------------------------------------------
N fertiliser:
- 9.70 10.16 4.29 6.25
+^a 10.65 9.45 5.62 5.52
LSD (P=0.05) 0.83 0.90
^a 41 kg N ha^-1^b Density of weed seedlings m-2 at 30 days after crop emergence transformed by SQRT(x + 0.5) ^c Density of weed inflorescences m-2 at crop maturity transformed by SQRT(x + 0.5) ^d Ba#1, Ba#2 = first barley crop and second consecutive barley crop in 3 year rotations, respectively ---------------------------------------------------------------------------
The density of grass weed inflorescences at maturity was affected by 2 main effects and 1 interaction component. On plots receiving annual application of 20 kg P ha^-1, total grass density (26 m^-2) was reduced (P ) from the level observed on 0 P plots (33 m^-2). Fertiliser P has previously been observed to reduce grass weed density in bread wheat in Ethiopia (Tanner et al., 1993), presumably by enhancing crop vigour and competitiveness. Grass weed density was higher (P) in the second successive barley crop after any precursor (34 m^-2) vis-a-vis the first barley crop (21 m^-2). However, there was also a significant interaction between first vs. second barley crop and fertiliser N (Table 7): the lowest density of grass weeds occurred in Ba#1-N. Previous research in Ethiopia has shown that fertiliser N exacerbates Avena fatua competitiveness in wheat, allowing the production of a higher density of wild oat panicles in the mixed canopy (Tanner et al., 1993). In the current study, approximately 90% of the grass weed inflorescences consisted of A. fatua panicles. CONCLUSION Interaction effects on weed density. Analysis of the 1994 data on density of broadleaf weed seedlings measured at 30 days after crop emergence indicated that the effects of rotation, Although barley GY responses to fertiliser nutrients were positive, particularly to applied P in the P-deficient soil of Bekoji and to fertiliser N in cereal rotations, crop rotation had the greatest impact on barley productivity. The effect of rotation with dicotyledonous crops interacted synergistically with fertiliser P to produce the highest yields of barley. The beneficial effects of dicot precursor crops on barley GY were largely dissipated in the second successive crop of barley. There was not a clear distinction between the effects of faba bean and rapeseed as precursor crops; this observation warrants further investigation. ACKNOWLEDGEMENT The trial discussed in this paper was supported financially by the Institute of Agricultural Research in Ethiopia and by the CIMMYT/CIDA Eastern Africa Cereals Programme. The authors wish to thank Tamirat Belay, Mekonnen Kassaye, and Workiye Tilahun for their participation in trial execution and data collection. REFERENCES Amanuel Gorfu, Tanner, D.G., Asefa Taa and Duga Debele. 1994. Observations on wheat and barley based cropping sequence trials conducted for eight years in southeastern Ethiopia. In: Developing Sustainable Wheat Production Systems. Tanner, D.G. (Ed.), pp. 261-280. CIMMYT, Addis Ababa, Ethiopia. Asefa Taa, Tanner, D.G. and Amanuel Gorfu. 1992. The effects of tillage practice on bread wheat in three different cropping sequences in Ethiopia. In: Seventh Regional Wheat Workshop for Eastern, Central and Southern Africa. Tanner, D.G. and Mwangi, W. (Eds.), pp. 376-386. CIMMYT, Nakuru, Kenya. Berhane Lakew, Hailu Gebre and Fekadu Alemayehu. 1996. Barley production and research. In: Barley Research in Ethiopia: Past Work and Future Prospects. Hailu Gebre and van Leur, J. (Eds.), pp. 1-8. IAR/ICARDA, Addis Ababa, Ethiopia. Cady, F.B. 1991. Experimental design and data management of rotation experiments. Agronomy Journal 83:50-56. Chilot Yirga, Hailu Beyene, Lemma Zewdie and Tanner, D.G. 1992. Farming systems of the Kulumsa area. In: Research With Farmers Lessons from Ethiopia. Franzel, S. and van Houten, H. (Eds.), pp. 145-157. CAB International, Wallingford, U.K. Cook, R.J. 1984. Root health: importance and relationship to farming practices, current technology and its role in sustainable agriculture. In: Organic Farming. Bezdicek, D.F. and Power, J.F. (Eds.), pp. 111-127. American Society of Agronomy, Madison, Wisconsin. Franzluebbers, A.J., Hons, F.M. and Saladino, V.A. 1995. Sorghum, wheat and soybean production as affected by long-term tillage, crop sequence and fertilization. Plant and Soil 173:55-65. Hailu Gebre, Amsal Tarekegne and Endale Asmare. 1989. Beneficial break crops for wheat production. Ethiopian Journal of Agricultural Science 11:15-24. Hargrove, W.L., Touchton, J.T. and Johnson, J.W. 1983. Previous crop influence on fertilizer nitrogen requirements for double-cropped wheat. Agronomy Journal 75:855-859. Heenan, D.P., Taylor, A.C. and Leys, A.R. 1990. The influence of tillage, stubble management and crop rotation on the persistence of great brome (Bromus diandrus Roth). Australian Journal of Experimental Agriculture 30:227-230. Higgs, R.L., Peterson, A.E. and Paulson, W.H. 1990. Crop rotations: sustainable and profitable. Journal of Soil and Water Conservation 45:68-70. Ladd, J.N. and Amato, M. 1985. Nitrogen cycling in legume-cereal rotations. In: Nitrogen Management in Farming Systems in Humid and Sub-Humid Tropics. Kang, B .T. and van der Helde, J. (Eds.), pp. 105-128. IBO/IITA, Ibadan, Nigeria. Lal, R. 1989. Conservation tillage for sustainable agriculture: Tropics versus temperate environments. Advances in Agronomy 42:85-197. Narwal, S.S., Malik, D.S. and Malik, R.S. 1983. Studies in multiple cropping. II. Effects of preceding grain legumes on the nitrogen requirement of wheat. Experimental Agriculture 19: 143-151. Patterson, H.D. 1965. The factorial combination of treatments in rotation experiments. Agricultural Science 65:171-182.
Preece, D.A. 1986. Some general principles of crop rotation experiments. Experimental Agriculture 22:187-198. Sinha, M.N., Hanpaiah, R. and Rai, R.K. 1983. Studies on phosphorus-use efficiency in cereal-cereal and legume-cereal rotations. Indian Journal of Agricultural Science 53:893-898. Tanner, D.G., Amanuel Gorfu and Asefa Taa. 1993. Fertiliser effects on sustainability in the wheat-based small-holder farming systems of Ethiopia. Field Crops Research 33:235-248. Taye Bekele. Yeshanew Ashagrie, Balesh Tulema and Girma Gebre Kidan. 1996. Soil fertility management in barley. In: Barley Research in Ethiopia: Past Work and Future Prospects. Hailu Gebre and van Leur, J. (Eds.), pp. 9299. IAR/ICARDA, Addis Ababa, Ethiopia. Zentner, R.P. and Campbell, C.A. 1988. First 18 years of a long-term crop rotation study in southwestern Saskatchewan - yields, grain protein and economic performance. Canadian Journal of Plant Science 68:1-21. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97020b.gif] [cs97020a.gif] |
| |||||||||

{kind=link}
{kind=link}