 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Genetic polymorphism of cassava within the Republic of Benin detected with RAPD markers N.J. Tonukari, G. Thottappilly, N. Q. Ng^1 and H.D. Mignouna
Biotechnology Research Unit a International Institute of Tropical
Agriculture, P.M.B. 5320, Ibadan, Nigeria (Received 14 February, 1997; accepted 21 August, 1997)
Code Number: CS97028
Sizes of Files:
Text: 24.2K
Graphics: Tables and line drawings (gif) - 37.2K
ABSTRACT A knowledge of the extent of genetic variation in a crop is a prerequisite for its improvement. In cassava (Manihot esculenta Crantz) such knowledge is lacking especially at the DNA level. The genetic variation among 28 cassava varieties collected from different locations in the Republic of Benin was investigated using random amplified polymorphic DNA (RAPD). A set of 10 random sequence 10-mer primers out of 100 that were screened detected polymorphisms and generated 78 amplified DNA bands. Based on an unweighted pair group method with arithmetic averages (UPGMA) cluster analysis, the accessions were categorised into 6 groups. The results, summarised in a dendrogram, show the genetic diversity among this cassava germplasm. Key Words: Cultivar identification, genetic diversity, Manihot esculenta, RAPDs RESUME La connaissance de l'importance de variation genetique chez une culture est necessaire pour son amelioration. Chez le manioc (Manihot esculenta Crantz), une telle connaissance manque surtout au niveau du DNA. La variation genetique parmi les 28 varietes du manioc recueillies de differentes localites du Benin etait etudiee en utilisant un polymorphe amplifie selectif du DNA (RAPD). Un ensemble de 10 amorces de 10-mer d'une selection sequentielle sur 100 qui avaient ete depistes ont detecte le polymorphisme et ont genere 78 bandes de DNA amplifie. Sur la base de l'analyse de faisceau de la methode d'un groupe de paire non-pondere avec les moyennes arithmetiques (UPGMA), les varietes etaient divisees en six groupes. Les resultats, resumes en dendogramme, montrent la variation genetique chez les differents types du plasme germinatif du manioc. Mots Cles: Identification varietale, variation genetique, Manihot esculenta, RAPDs Introduction Cassava (Manihot esculenta Crantz) is a major carbohydrate food for millions of people world-wide, and one of the most important crops in the tropics. In tropical Africa, cassava is cultivated mainly for its storage root which is the single-most important source of calories in the diet (Cock, 1982; Hahn, 1989). Efforts are being made to develop cassava varieties with high and stable yield across different agro-ecologies. Success in these efforts depends on the availability of an extensive level of genetic variability. Some amount of variation necessary for a genetic improvement programme of cassava cultivars already exist within the several germplasm collections from different parts of Africa (Beeching et al., 1993). However, there is no adequate information on the extent of genetic variation available in this crop. Genetic variation studies using isozyme analysis has been attempted in cassava (Hussain et al., 1987; Ramirez et al., 1987; Lefevre and Charrier, 1993), but the results showed low level of polymorphism. Besides, isozyme patterns were found to be influenced by stage of plant development as well as the part of the plant from which sample was taken. There are other molecular techniques that can detect variability within and among several crop species. Two of these widely used techniques are restriction fragment length polymorphisms (RFLPs) (Helentjaris et al., 1986) and specific amplification of polymorphic DNA fragments by PCR (Weining and Langridge, 1991). Using specifically amplified cassava cDNA as probes, Beeching et al. (1993) detected RFLP in Manihot germplasm, assembled from different parts of the world. Their study revealed significant levels of polymorphism both within and between the species. Bertram and Schaal (1993) have also used RFLP to study the phylogeny of Manihot and the evolution of cassava. This technique, unlike isozyme analysis, detects more poly-morphisms and has the advantage of being consistent between tissues. Nevertheless, the intense labour and time inputs (Kazan et al., 1993) and the requirements of useful probes most times render RFLP analysis cumbersome. The polymerase chain reaction (PCR) allows for the detection of random amplified polymorphic DNAs (RAPDs) using arbitrarily designed short primers to amplify DNA (Welsh and McCleland, 1990, 1991; Williams et al., 1990). RAPD requires no previous sequence information or probe identity for the fingerprinting of cultivars. Moreover, the DNA fragment electrophoretic patterns generated are able to discriminate between interspecific and intraspecific variation in several plant species (Caetano-Anolles et al., 1991; Hu and Quiros, 1991; Hadrys et al., 1992). Marmey et al. (1994) have successfully detected RAPDs using 8 primers, among 3 species of Manihot and 19 cultivars of M. esculenta. However, these cultivars and species were assembled from many countries in Africa, India and South America. Our aim was to determine whether RAPDs are useful for cultivar discrimination and identification in cassava populations collected from a small geographical region. In this study, PCR-RAPD was evaluated for its ability to detect polymor-phisms within 28 germplasm accessions collected from the Republic of Benin in West Africa. MATERIALS AND METHODS
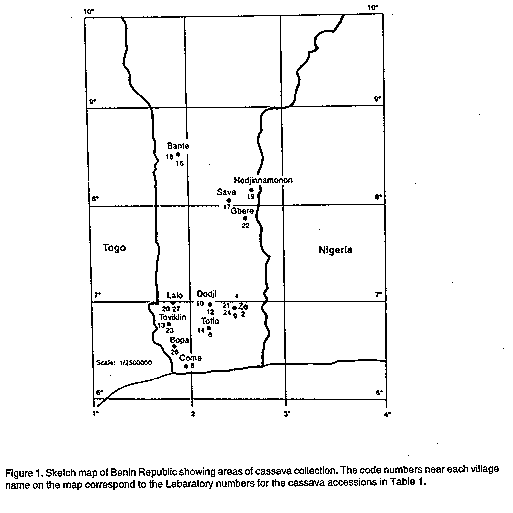
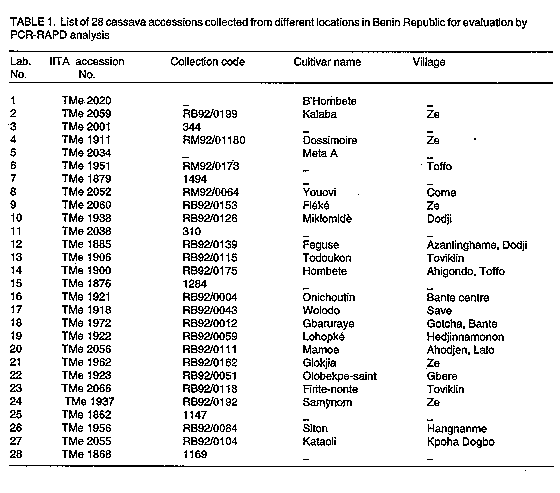
Plant material. The cassava clones used in this study were collected from farmers' fields in at least 11 locations in the Republic of Benin (Fig. 1, Table 1). Most of the accessions came from the southern part of the country (between latitudes 6 degrees N and 9 degrees N) where cassava is mostly cultivated. The plants were grown from stem cuttings and maintained in the screenhouse of the Germplasm Resources Unit at the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria. DNA was extracted from plants grown in pots placed in the screenhouse. DNA isolation. The procedure used for total genomic DNA extraction was that of Saghai-Maroof et al. (1984) with some modifications. Young leaf tissue from single plants of each accession was ground to a fine powder in liquid nitrogen. Approximately 100 mg of the powdered tissue was suspended in 600 ul of 2 x CTAB buffer (50 mM Tris, pH 8.0; 0.7 M NaCl; 10 mM EDTA; 1% hexadecyltrimethylammonium bromide; 0.1% 2-mercaptoethanol) and incubated at 55 C for 30 min. DNA was purified by two extractions with chloroform:isoamyl alcohol (24:1) and precipitated with ice-cold absolute ethanol. After washing with 70% ethanol, the DNA was dried and resuspended in 100 ul TE (10 mM Tris-HCl, 1 mM EDTA, pH 8.0). DNA concentration was measured using a DU-65 UV spectrophotometer (Beckman Instruments Inc., Fullerton CA, USA) at 260 nm. To check for degradation of the DNA, the samples were loaded onto a 1.0% agarose gel in 0.5 x TBE (45 mM Tris-borate, 1 mM EDTA, pH 8.0) and electrophoresed.
PCR and electrophoresis. DNA primers tested in this study were purchased from Operon Technologies (Alameda, CA ,USA) and each was 10 nucleotides long. Two concentrations of each template DNA (24 ng and 96 ng per reaction) were used to test reproducibility and eliminate sporadic amplification products from the analysis. Amplifications were performed in 12 l reaction mixture consisting of genomic DNA, 1 x reaction buffer (Promega Corporation, Madison WI, USA), 100 mM each of dATP, dCTP, dGTP and dTTP, 0.2 uM primer, 2.5 uM MgCl2 and 1 U of Taq polymerase (Boehringer Mannheim GmbH, Germany). A single primer was used in each reaction. The reaction mixture was overlaid with 50 ul of mineral oil to prevent evaporation. Amplification was performed in a thermowell microtiter plate using a PTC-100^a programmable Thermal Controller (MJ Research Inc., Watertown, MA, USA). The cycling programme involved an initial denaturation at 94 C for 3 minutes and followed by 40 cycles of 94 C for 1 minute, 35 C for 1 minute and 72 C for 2 minutes; and a final extension at 72 C for 10 minutes. Amplification products were maintained at 4 C until electrophoresed. The reaction products were resolved by electrophoresis in a 1.4 % agarose gel in 0.5 x TBE buffer at 150 V for 2.5 hours. A 1-kb ladder (Life Technologies, Gaithersburg, MD, USA) was included as size marker. Gels were visualised by staining with ethidium bromide solution (0.5 ug ml^-1) and banding patterns were photographed over UV light using a red filter. Data analysis. RAPD markers that were consistently reproduced in at least two replicate PCR reactions were further analysed. Fragment sizes of RAPD were estimated from the gel photograph by comparison with the 1-kb ladder marker. From the whole set of reproducible RAPD bands generated in the 28 cassava clones by the different primers, bands were scored as present (1) or absent (0). A data matrix with fragments in columns and accessions in lines was assembled from these scores. Only the polymorphic markers were included in the data set. Arbitrarily primed PCR characters were analysed using Genstat 5 Release 3.1 (Lawes Agricultural Trust, Rothamsted Experimental Station, Harpenden, Hertfordshire, U.K.). A similarity/distance matrix was developed using Jaccard (1901) similarity coefficient (Sneath and Sokal, 1973), which does not consider the joint absence of a marker as an indication of similarity. The similarity index of Jaccard between plants i and j is given by:
RESULTS One hundred random oligonucleotide primers (Operon Technologies sets A, B, D, E and O) were evaluated for their ability to prime PCR amplification of cassava genomic DNA. In a preliminary survey, genomic DNA of two of the accessions, TMe 1900 and TMe 1921 collected from Ahingodo, Toffo and Bante, respectively (see Fig. 1), were used as templates. Eighty one of the primers did not amplify the two DNA templates or resulted in only limited amplifications that were visualised on gels as faint bands or smears. The remaining 19 primers were able to amplify the genomic DNA, giving reproducible RAPD amplification patterns with individual fragments that stained intensely. Ten of these primers (Table 2) were chosen for the whole experiment and used to amplify the DNA of the various accessions. Examples of typical PCR amplification products are shown in Figure 2.
The 10 informative oligonucleotide primers detected polymorphisms among the 28 cassava accessions. The primers revealed a total of 78 clear and easily scorable bands. The number of bands per primer ranged from 6 to 12. The size of DNA bands that were produced in the PCR reactions ranged from 300 to 2100 bp, but most of the bands were between 500 and 1000 bp. None of the primers was specific for any of the accessions. However, primers OPE-03 and OPE-04 produced a 480 bp and 1200 bp bands, respectively, in TMe 1862 and TMe 2052 only. Also a 920 bp band was generated in TMe 1876 and TMe 2052 only by primer OPF-01, while a 900 bp band generated by OPO-20 was specific only to TMe 1918 and TMe 1921. Examples of polymorphic bands used for discrimination among the accessions are shown in Figure 2. Only twenty five (32%) of the 78 bands were poly-morphic among the cassava accessions. Data from the scores were subjected to UPGMA analysis. Jaccard similarity coefficient ranged from 100% for the most closely related accessions to approximately 40% for those most distantly related, with a mean of 69%. With our PCR conditions, TMe 1876 showed the highest polymorphic bands while TME 1906 showed the least. None of the primers could provide enough markers to discriminate between all the cassava populations. Identical RAPD patterns (100% Jaccard similarity coefficient) using the ten primers were observed for four pairs of the accessions; TMe 1900 and TMe 2055, TMe 1918 and TMe 1921, TMe 1972 and TMe 2056, and TMe 2059 and TMe 2066. Also, TMe 1937 and TMe 1951 as well as TMe 1906 and TMe 1911 has 95% and 94% similarity coefficients, respectively. The similarities between accession TMe 2052 and accessions TMe 1906, TMe 1900 and TMe 2001 were 40%, 42% and 42%, respe-ctively. Based on the average cluster analysis, the accessions formed 6 groups at the 70% Jaccard similarity coefficient level (Fig. 3).
DISCUSSION PCR-RAPD technique provided means for genetic analysis of cassava cultivars collected from farmers' fields in a small geographical region (Republic of Benin, 116,347 km^2). This methodology was developed to identify Manihot species and 19 cassava cultivars assembled from many countries in Africa, India and South America (Marmey et al., 1994). The use of RAPD analysis also revealed extensive polymorphism in other species (Carlson et al., 1991; Hu & Quiros, 1991; Welsh et al., 1991; Reiter et al., 1992; Wolff & Rijn, 1993), showing that RAPD technique is able to detect diversity at the intraspecific level. Using the RAPD patterns generated by the 10 random primers with G+C content ranging from 60 to 70%, it was possible to detect genomic polymorphisms in Manihot esculenta. Only a combination of RAPD patterns allowed the unambiguous characterisation of the cassava accessions. Although the cluster analysis assigned the accessions into 6 groups, none of these, except TMe 2052, were classified according to geographical origin. TMe 2052, the only accession collected from Come (Fig. 1) differed considerably from all the others, having only 40 and 42% similarities with TMe 1900 and TMe 2001, respectively. Similarly, the dendrogram generated by Marmey et al. (1994) using 8 primers and 19 cassava accessions did not provide a strong group structure based on geographical origin. Nevertheless, their study indicated genetic diversity within the population studied with only 2 accession pairs having zero genetic distances. Moreover, the accession pairs having 100% Jaccard similarity coefficients [TMe 2059 (Ze) and TMe 2066 (Toviklin); TMe 1918 (Save) and TMe 1921 (Bante); TMe 1900 (Toffo) and TMe 2055 (Lalo); and TMe 1972 (Bante) and TMe 2056 (Lalo)] were also found to have originated from different locations as indicated after the accession numbers. Although they were collected from relatively distant regions (Fig. 1), our RAPD analysis indicated that these accessions were closely related based on their similarity coefficients. This situation is not unusual (Beeching et al., 1993) particularly for an introduced crop such as cassava, which is predominantly vegetatively propagated. The basic similarities also observed among the cassava accessions used in this study was also not unexpected due to the small geographical region from which the plants were collected. Cultivar identification is an important issue for breeders and germplasm managers. The application of RAPDs seems very valuable in this regard. Using RAPD to determine the introgression of desirable traits such as disease resistance should therefore allow breeders to select related or unrelated parental germplasm material to maximize variability in their breeding programs as has been pointed out by similar studies in cassava (Chvez, 1990; Beeching et al., 1993; Marmey et al., 1994). There are over 2000 accessions of cassava germplasm currently maintained at IITA. In collaboration with national scientists and agricultural authorities in Africa, the Institute intends to continue to assemble the germplasm collection in Africa. The methodolgy of PCR-RAPD established in this study with additional assessment of other primers and techniques will be intensified for the investigation of diversity and detection of duplicate germplasm samples. This will enhance the efficiency of germplasm management and the use of cassava germplasm. A more extensive sampling of populations from different parts of tropical Africa would definitely reveal a greater genetic variability. In summary, this study reveals the genetic diversity of cassava accessions collected from the southern part of the Republic of Benin. We suggest that RAPD, or other appropriate genetic markers, be used whenever possible in evaluating collections of cassava cultivars. ACKNOWLEDGEMENT We would like to acknowledge the collaboration of Messr Sanni Abou and Maroyo Norbert of the National Agricultural Research Station, Niaouli, Republic of Benin. The authors also wish to thank J. Ikea, O. G. Omitogun, I. Ingelbrecht, A. Pellegrineshi, B. Ubi, C. A. Fatokun, R. Terauchi, R. Ortiz, A. Dixon, R. Asiedu and J. Crouch for their critical reading of the manuscript and useful comments. REFERENCES Beeching, J.R., Marmey, P., Gavalda, M-C., Noirot, M., Hayso, H.R., Hughes, M.A. and Charrier, A. 1993. An assessment of genetic diversity within a collection of cassava (Manihot esculenta Crantz) germplasm using molecular markers. Annals of Botany 72:515-520. Betram, R.B. and Schaal, B.A. 1993. Phylogeny of Manihot and the evalution of cassava. CBN- Newsletter 1:4-6. Caetano-Anolles, G., Bassam, B.J. and Cresshorff, P.M. 1991. DNA amplification fingerprinting using very short arbitrary oligonucleotide primers. Bio/Technology 9:553-557. Carlson, J.E., Tulsieram, L.K., Glaubitz, J.C., Luk, V.W.K., Kauffeldt, C. and Rutledge, R. 1991. Segregation of random amplified DNA markers in F1 progeny of conifers. Theoritical Applied Genetics 83:194-200. Chavez, R. 1990. Wild Manihot species: a valuable resource. Cassava Newsletter 14:2-5. Cock, J.H. 1982. Cassava: a basic energy source in the tropics. Science 218:755-762. Hadrys, H., Balick, M. and Schierwater, B. 1992. Applications of random amplified poly-morphic DNA (RAPD) in molecular ecology. Molecular Ecology 1:55-63. Hahn, S.K. 1989. An overview of African traditional cassava processing and utilization. Outlook in Agriculture 81:110-118. Halward, T., Stalker, T., Larue, E. and Kochert, G. 1992. Use of single-primer DNA amplifications in genetic studies of peanut (Arachis hypogaea L.). Plant Molecular Biology 18:315-325. Helentjaris, T., Slocum, M., Schaefer, A. and Nienhuis, J. 1986. Construction of genetic linkage maps in maize and tomato using restriction fragment length polymorphisms. Theoritical Applied Genetics 72:761-769. Hu, J. and Quiros, C.F. 1991. Identification of broccoli and cauliflower cultivars with RAPD markers. Plant Cell Report 10:505-511. Hussain, A., Bushuk, W., Ramirez, H. and Roca, W. 1987. Identification of cassava (Manihot esulenta Crantz) cultivars by electrophoretic patterns of esterases enzymes. Seed Science Technology 15:19-21. Jaccard, P. 1901. Etude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaudoise Sci. Nat. 37:547-579. Kazan, K., Manners, J.M. and Cameron, D.F. 1993. Inheritance of random amplified polymorphic DNA markers in an interspecific cross in the genus Stylosanthes. Genome 36:51-56. Lefevre, F. and Charrier, A. 1993. Isozyme diversity within African Manihot germplasm. Euphytica 66:73-80. Liu, Z.-W., Jarret, R.L., Duncan, R.R. and Kresovich, S. 1994. Genetic relationships and variation of ecotypes of seashore paspalum (Paspalum vaginatum) determined by random amplified polymorphic DNA markers. Genome 37:1011-1017. Mailer, R.J., Scarth, R. and Fristensky, B. 1994. Discrimination among cultivars of rapeseed (Brassica napus L.) using DNA poly-morphisms amplified from arbitrary primers. Theoritical Applied Genetics 87:697-704. Marmey, P., Beeching, J.R., Hamon, S. and Charrier, A. 1994. Evaluation of cassava (Manihot esculenta Crantz) germplasm using RAPD markers. Euphytica 74:203-209. Ramirez, H., Hussain, H., Roca, W. and Bushuk, W. 1987. Isozyme electrophoregrams of sixteen enzymes in five tissues of cassava (Manihot esculenta Crantz) varieties. Euphytica 36:39-48. Reiter, R.S., Williams, J.G.K., Feldmann, K.A., Rafalski, J.A., Tingey, S.V. and Scolnik, P.A. 1992. Global and local genome mapping in Arabidopsis thaliana by using recombinant inbred lines and random amplified polymorphic DNAs. Proceedings National Academy of Science U.S.A 89:1477-1481. Saghai-Maroof, M.A., Soliman, K.M., Jorgensen R.A. and Allard, R.W. 1984. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proceedings National Academy of Science U.S.A 81:8014-8018. Sneath, P.H. and Sokal, R.R. 1973. Numerical Taxonomy. The Principles and Practice of Numerical Classification. W. H. Freeman and Co., San Francisco, USA. Tao, Y., Manners, J.M., Ludlow, M.M. and Henzell, R.G. 1993. DNA polymorphisms in grain sorghum [Sorghum bicolor (L.) Moench]. Theoritical Applied Genetics 86:679-688. Weining, S. and Landridge, P. 1991. Identification and mapping of polymorphisms in cereals based on the polymerase chain reaction. Theoritical Applied Genetics 82:209-216. Welsh, J. and McClelland, M. 1990. Finger-printing genomes using PCR with arbitrary primers. Nucleic Acids Research 18:7213-7218. Welsh, J. and McClelland, M. 1991. Genomic fingerprinting with arbitirary primed PCR and a matrix of pairwise combination of primers. Nucleic Acids Research 19:5275-5279. Welsh, J., Honeycutt, R.J., McClelland, M. & Sobral, B.W.S. 1991. Parental determination in maize hybrids using the arbitrary primed polymerase chain reaction (AP-PCR). Theoritical Applied Genetics 82:473-476. Williams, J.G.K., Kubelik, A.R., Livak, K.I., Rafalski, J.A. and Tingey, S.V. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research 18:6531-6535. Wolff, K. and Rijn, J.P. 1993. Rapid detection of genetic variability in chrysanthemum (Dendrathema grandiflora Tzvelev) using random primers. Heredity 71:335-341. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97028e.gif] [cs97028c.gif] [cs97028d.gif] [cs97028b.gif] [cs97028a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}