 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
In vitro shoot multiplication of Eucalyptus grandis F. WACHIRA Tea Research Foundation of Kenya, P. O. Box 820, Kericho, Kenya (Received 24 September, 1996; accepted 8 July, 1997)
Code Number: CS97030
Sizes of Files:
Text: 33.3K
Graphics: Tables and line drawings (gif) - 51.8K
ABSTRACT In vitro shoot multiplication was induced from sections of orthotropic shoot sprout regrowth of 25 year old pruned Eucalyptus grandis using a regeneration medium supplemented with several levels of different plant growth regulator combinations, different carbon sources, and different pH at time of inoculation. Optimal shoot regeneration was obtained in Murashige and Skoog medium supplement with 0.4mg.l^-1 BAP and 1.0mg.l^-1 IBA or 0.2mg.l^-1 BAP and 1.0mg.l^-1 NAA and 3% (w/v) sucrose. An 8-week subculture period was found to be suitable. Both NAA and IBA decreased the shelf life of E. grandis shoot multiplication cultures by encouraging callus growth and proliferation. Sucrose at 30g.l^-1 gave superior growth compared to the other three carbon sources tested, i.e., maltose, glucose and fructose. The pH of the medium affected growth of the shoot cultures with a pH of 5.8 giving better growth. Key Words: Eucalyptus, plant growth regulators, rooting RESUME La reproduction des pousses in vitro etait induite des zones orthotropiques du bourgeon de la pousse en etat de recroissance d'un eucalyptus taille et age 25 ans, en utilisant une regeneration d'un media supplemente avec plusieurs differentes combinaisons regulatrices de croissance des plantes, des sources carboniques differentes, et un PH different au moment de l'inoculation. La regeneration optimale de la pousse etait obtenue dans le media de murashige et Skoog supplee avec 0.4 mg l^-1 IBA ou 0.2 mg l^-1 IBA ou 0.2 mg l^-1 BAP et 1.0 mg l^-1 NAA et 3% (W/V) du sucrose. Une periode sous-culturelle de 8 semaines etait jugee convenable. NAA et IBA reduisaient la duree de vie de culture des pousses de E. grandis en encourageant la croissance et la proliferation des calles. Le sucrose administre 30 g/l^-1 a donne une croissance elevee par rapport aux autres sources carboniques testees, telles que le maltose, le glucose et le fructose. Le PH du media a affecte la croissance des cultures de pousses avec de proportions de 5.8 de PH, assurant par consequent la meilleure croissance. Mots Cles: Eucalyptus, regulateurs de croissance, encracinement INTRODUCTION Despite the fact that eucalypts are bisexual with fertile male and female organs in the same flower, and to some degree are self-compatible, they are predominantly outbreeding (Hodgson, 1976a) with intra-floral self-pollination and self-fertilisation diminished by protandry (Hodgson, 1976b; Wyk, 1977) and a probable gene controlled incompatibility system (Hodgson, 1974; Eldridge, 1976). Seed propagated plants therefore exhibit great variability in yield potential and other genetic characters. Several attempts have been made to clone elite trees via cuttings and in vitro techniques. Mass production, via cuttings has been most successful when adventitious or epicormic shoots from 3-5 year-old trees are used. Up to 600 cuttings can be propagated from one single tree per year (FAO, 1981). Chaperon (1978) was the first to achieve mass propagation by cuttings on a semi-industrial scale. Several attempts have also been made to mass propagate eucalypts via in vitro culture techniques (Durzan and Campbell, 1974; Fossard et al., 1974; Konar and Nagmani, 1974; Burger, 1987; Curir et al., 1987; Damiano et al., 1987).
The first forest trees that were clonally propagated in vitro were juvenile aspen (Winton, 1970). Up to 10,000 plantlets were produced from a single seedling in a year (Nadgir et al., 1984). One of the major limitations in tree micropropagation has, however, remained the failure of explants from mature trees to undergo organogenesis. Little is known of the physiological and genetic mechanisms that control the differences between the juvenile and mature physiological state of cells, tissues or the plantlet, though it has been shown by Paton et al. (1970) that mature leaves of Eucalyptus grandis produce a chemical that inhibits root development. The role of adult leaves in induction or maintenance of a mature behaviour of the meristem was also demonstrated by Doorenbos (1965) in Hedera helix.
In large scale clonal propagation programmes, the number of juvenile branches can be increased by several means. In vitro culture of orthotropic shoots produced by repeated pruning (hedging) has been successfully achieved in Hevea (Mascarenhas et al., 1982) and E. sideroxylon (Burger, 1987). Juvenility has also been routinely induced by spraying growth regulators at the base of the trunk.
This has been done in E. ficifolia using 6-benzyl aminopurine (BAP) (Mozalewski and Hackett, 1979) and in Hedera (ivy) using gibberellin (Robbins, 1957). Repeated subculture of tissues excised from crowns has also been used to induce juvenility (Creswell and Nitch, 1975). Using this technique, thirty-year-old frost resistant trees of E. gunnii (Boulay and Franclet, 1983) and mature trees of E. citridora (Gupta et al., 1981) were successfully propagated in vitro.
Most shoot multiplication cultures in eucalypts have been initiated on modified Murashige and Skoog (1962) - MS medium often incorporating 6-benzylaminopurine (BAP),- napathalene acetic acid (NAA), indole-3-butyric acid (lBA) and kinetin (K) either singly or in combination as plant growth stimulators (Hartney, 1982; Burger, 1987; Jalil and Gavinlertvatana, 1989). Other less commonly used media have also been successfully used (Curir et al., 1987; Damiano et al., 1987).
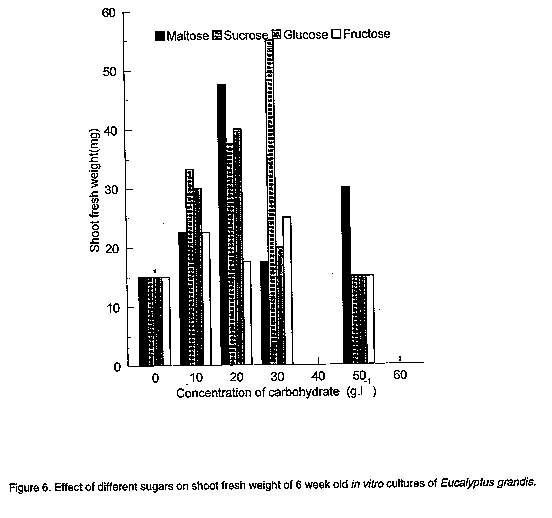
The present study evaluates the reaction of shoot cultures of a mature Kenyan stand of E. grndis to several culture factors which include plant growth regulator combinations, carbon source and pH of medium at time of culture. MATERIALS AND METHODS Nodal explant tissues (measuring about 1.5 cm) were obtained from orthotropic shoots of sprout regrowth from 25 year old pruned trees and used as propagules. Two weeks prior to collecting the explants, the tree stumps had been sprayed with a copper based fungicide, Kocide 101 at a rate of 7 mg.l^-1 in water per stump. The explants were cultured under aseptic conditions on semi-solid Murashige and Skoog (1962)-MS medium (pH 5.8 before autoclaving at 121 C for 15 minutes) containing various plant growth regulators. The media contained Fe-EDTA and the MS-micronutrients plus 0.5mg. l^-1 thiamine-HCl, 0.5mg.l^-1 pyridoxine-HCl, O.05mg.l^-1 nicotinic acid, 100mg.l^-1 m-inositol, 8g.l^-1 Difco bacto-agar and 30g.l^-1 sucrose unless stated otherwise. In each case, 10 ml of the medium was used in a 23 ml culture jar bottle. Before culturing, the explant tissues were soaked in distilled water for one hour then pretreated in 2% polyvinyl pyrollidone (PVP) for 30 minutes, surface sterilised in 2% mercuric chloride for 15 minutes and subsequently washed three times in autoclaved double distilled water. All cultures were incubated at 5 C in darkness for 48 hours before being exposed to light. The cultures were then maintained at 24+/-2 C daytime and 18+/-2 C night time temperatures and a 13-hour photoperiod at a flux density of 2,800 lux. Unless stated otherwise, the variables recorded after 8 weeks culture were the number of usable (>5mm) shoots per responsive explant. Each treatment in all the experiments described, was applied to 8 explants and was repeated 3 times. All data were subjected to analysis of variance. Effect of BAP combined with NAA on shoot regeneration and multiplication. Since auxin and cytokinin concentrations are critical for establishing a plant regeneration system, an experiment was initiated using nodal explants which were cultured in MS medium in a factorial arrangement of 0, 0.2, 0.4, 1.0, 2.0, 4.0 and 8.0mg.l^-1 BAP and O, 0.4, 1.0, 2.0mg.l^-1 NAA concentrations. Influence of BAP combined with IBA on shoot regeneration and multiplication. A similar experiment as that described in 1 above was initiated but with NAA substituted with IBA. The same levels of auxin and cytokinin as described earlier, were tested. Influence of NAA and IBA separately on shoot regeneration frequency. The effect of auxins (NAA and IBA separately) on shoot regeneration frequency with time was evaluated in two experiments in which 4 levels of auxin (0, 0.4, 1.0 and 2.0 mg.l^-1) were tested in replicated experiments. Shoot numbers were scored weekly during the experimental period. Effect of the initial pH of the medium at time of inoculation on in vitro shoot growth. To investigate the effect of the initial pH of the medium on the in vitro growth of Eucalyptus grandis, nodal explants each weighing 10 mg were handled as described in the earlier experiments and cultured on MS medium without phytohormones but fortified with 30g.l^-1 sucrose. The initial pH of the medium had been adjusted to 3.8, 4.8, 5.8, 6.8 and 7.0 respectively, before autoclaving. Shoots produced were then scored for their fresh weight after 4 weeks growth in culture. Dry weight was established by oven-drying the shoots at 90 C for 48 hours. The effect of type and concentration of carbon/energy source on shoot growth. The effect of exogenously supplied carbohydrates on shoot growth was evaluated in an experiment in which four different carbon sources (maltose, sucrose, glucose and fructose) were tested in MS medium (pH 5.8 before autoclaving) without plant growth regulators. Nodal explants each weighing 10mg were excised, inoculated in medium supplemented with the four different sugars each at 5 levels (0, 10, 20, 30 and 50g.l^-1). After 6 weeks growth in culture, the resulting shoots were removed, cleaned and their fresh weight established. Rooting experiments. The induction of roots from in vitro cultured shoots was attempted in one and half strength MS medium supplemented with nine different concentrations of three auxins (IBA, NAA and IAA) as shown in Table 1, Table 2 and Table 3. All other culture conditions were as described above.
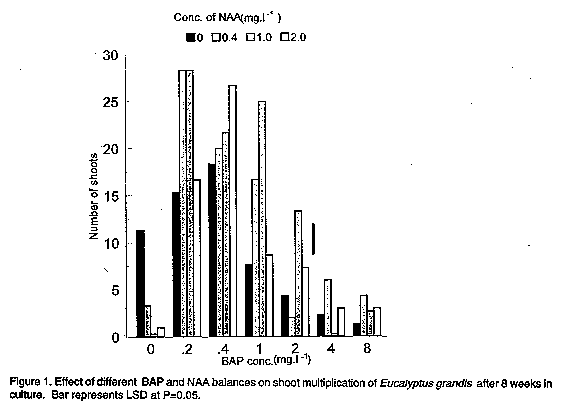
RESULTS AND DISCUSSION The initial soaking of the explants in distilled water, coupled with pretreatment in PVP and initial incubation at 5 C in darkness, markedly reduced the accumulation of oxidised polyphenolic compounds which are often associated with media browning and subsequent explant death. Cresswell and Nitsch (1975), similarly overcame the problem of medium browning in in vitro cultures of E. grandis by soaking sterilised explants in water for at least 2 hours and by keeping the cultures in darkness for the first 7 days. In general, the stump sprout nodal explants used in the experiments described, responded satisfactorily to in vitro culture and shoot buds were first observed within 1 to 2 weeks of exposure to light and room temperature. Successful use of coppice regrowth explants in micropropagation has been reported in E. sideroxylon (Burger, 1987), and positional effects have been shown to influence in vitro and vegetative propagation in Eucalyptus (Durand-Cresswell et al., 1982) as well as in some other trees (Rouland, 1973). Explants obtained from lower branches especially in positions near the trunk, are therefore preferred (Bonga, 1987). Microcuttings with leaf nodes excised from stems or side shoots of young seedlings have also been successfully used as explants in the micro-propagation of E. gunnii (Curir et al., 1987). Reports on the successful micropropagation of mature Eucalyptus trees have also been made by use of tissues excised from crowns (Gupta et al., 1981; Boulay and Franclet, 1983) and lateral buds (Jalil and Gavinlertvatana, 1989). The range of cytokinin concentrations allowing bud and shoot regeneration was large, i.e., from 0 to at least 3.0mg.l^-1. In experiment 1, optimum shoot regeneration was achieved at 0.2mg.l^-1 BAP and 0.4mg.l^-1 or 1.0mg.l^-1 NAA (Fig.1). An average of 28 shoots were produced per nodal explant after 8 weeks in culture ( Plate 1a). Shoot regeneration increased quadratically with increase in BAP concentration. BAP-NAA interactions on shoot regeneration were statistically significant (P<0.01). In vitro culture studies of Eucalyptus have demonstrated that shoot multiplication rates depend upon the species, whether or not the propagules are juvenile or mature and the individual genotype of the tree (Hartney, 1981; McComb and Bennett, 1986). Curir et al. (1987) reported the production of 5-20 shoots from E. gunnii microcuttings cultured in modified Fossardedium with BAP. Hartney (1983) also demonstrated the viability of micro-propagation systems using 21 Eucalyptus species. In his study, which specifically demonstrated the immediate commercial value of micropropagation for E. caesia and E. macrocarpa, shoot multi-plication was achieved at one and half strength MS, whilst rooting was achieved at one and half strength MS media.
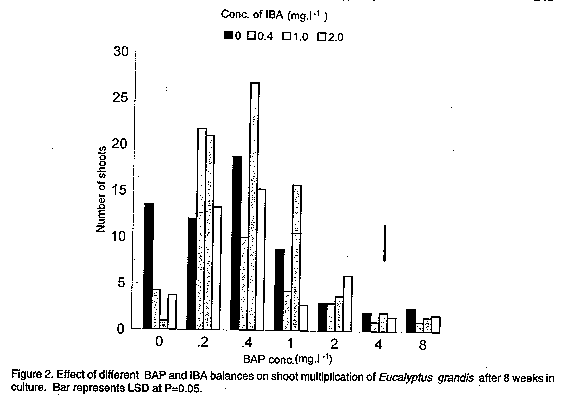
In experiment 2, the highest shoot harvest was obtained at 0.4mg.l^-1 BAP and 1.0mg.l^-1 IBA (Fig. 2). Quadratic responses with increase in BAP concentration were scored for shoot numbers in all the IBA treatments. Without BAP, addition of IBA severely suppressed shoot regeneration. Above 2.0mg.l^-1 BAP, induction and growth of bud primordia to form shoots was suppressed irrespective of IBA concentration. BAP-IBA interactions were significant (P<0.01). To check and minimise the growth of basal callus, an 8-week transfer (subculture) period was found to be suitable for all shoot multiplication cultures. With an 8-week culture period, a constant supply of shoots can be regularly harvested for further plantlet regeneration.
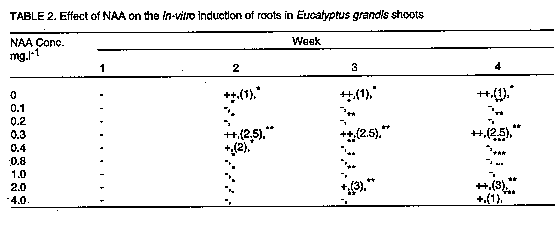
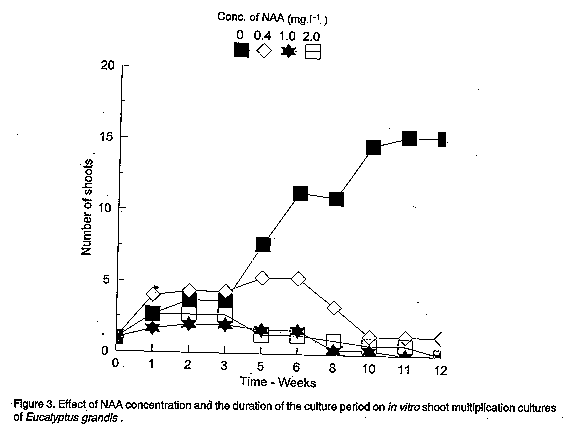
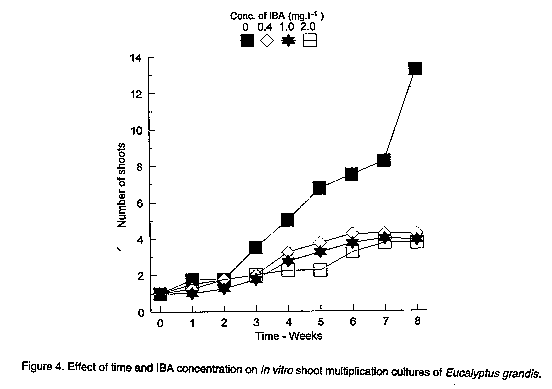
In the absence of cytokinin in the culture medium, NAA and IBA decreased the shelf life of the cultures by promoting callus development at the expense of shoot multiplication. Increasing concentration of either NAA (Fig. 3) or IBA (Fig. 4) decreased the number of harvestable shoots and in some cases, cultures were almost completely overgrown with callus tissue ( Plate 1b). Similar observations have been made in the micropropagation of juvenile tissues of Pinus nigra by Kolevska-Pletikapic et al. (1983). Some shoot cultures also rooted sporadically (Plate 1c).
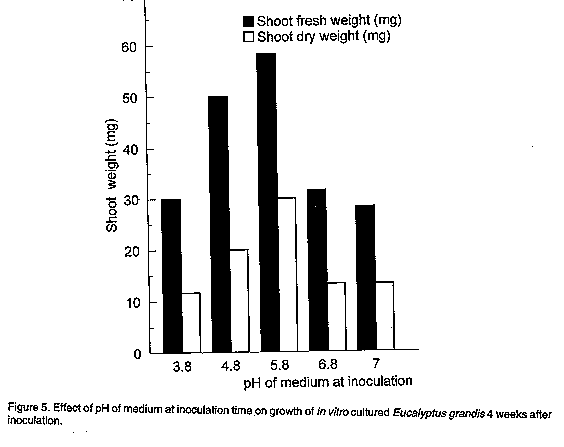
The medium pH at inoculation time insignificantly affected the size of harvestable shoots as measured by fresh and dry weights. An initial pH of 5.8 before autoclaving gave marginally heavier shoots Fig. 5). The operating pH during the growth period in culture was not measured in this experiment, though it was expected that there would be a shift following autoclaving and inoculation as has been demonstrated in other studies (Erickson, 1965; Durzan and Chalupa, 1974; Martin and Rose, 1976). According to Minocha (1987), cell growth in culture is often not significantly affected within a broad range (4.0 - 7.0) of initial medium pH. Some exceptions to this have been reported. For example, an initial pH of 7.5 (6.8 after autoclaving) was found optimal for the growth of Nicotiana glutinosa, N. tabacum and some Populus hybrids (Matsumoto et al., 1972). A pH of 5.4 was found by Wetherell and Dougal (1976) to be optimal for growth and embryogenesis in carrot cultures.
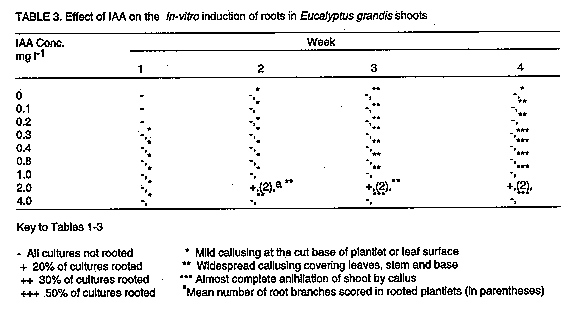
Although carbohydrates are of prime importance for in vitro organogenesis, carbon metabolism in vitro is still not clearly understood (Kozai, 1991). However, it is well established that carbohydrate requirements depend upon culture stage and differ among species (Thompson and Thorpe, 1987). The effect of type and concentration of exogenously supplied carbohydrate on shoot fresh weight of E. grandis is presented in Figure 6. All carbohydrate sources tested gave quadratic responses to concentration with optima at 20 or 30g.l^-1. Carbohydrate concentration modifies the osmotic strength of the medium thereby strongly affecting osmoregulation (Thompson and Thorpe, 1987). Osmoregulation and growth are inextricably linked and a high osmotic strength of the medium may result in reduced plant growth, height, and fresh weight. The best response in these experiments was recorded for sucrose at 30g.l^-1 which is within the widely used range of 20-40g.l^-1 (Thorpe and Patel, 1984). Fructose gave the poorest response. Both glucose and maltose had their response maxima at 20g.l^-1. Though several tissue culture studies have shown sucrose to be the best carbon and energy source (Street, 1969; Von Arnold and Eriksson, 1981; Thorpe, 1982), other carbohydrates have also been used successfully. Glucose, for example, was demonstrated to be an efficient carbon source for Cork Oak culture (Romano et al., 1995), Alnus (Welander et al., 1989), Potentilla fructicosa and Ficus lyrata (Wainwright and Scrace, 1989). Sugar alcohols have also been shown to support in vitro growth in apple and related species (Pua and Chong, 1984; Marino et al., 1993). The poor response to fructose noted in this study may indicate that it is not efficiently metabolised by E. grandis. Autoclaved fructose has been shown to have inhibitory effects on in vitro cultures of some species e.g. Cork Oak (Romano et al., 1995). Contradictory results to these have, however, been obtained for other species, e.g. mulberry (Oka and Ohyama, 1982; 1986), Malus (Welander et al., 198; Moncousin et al., 1992), Syringa and Alnus (Welander et al., 1989), etc. Such differences in species responses to autoclaved fructose is thought to be due to differential species sensitivity to degradation products formed as a result of autoclaving such as furfural and hydroxymethylfurfural (Hsiao and Bornman, 1989). The suggestion that the carbohydrate source could influence the type of organ differentiation from the primary explant (Kikuta and Okazawa, 1984) was not demonstrated in the present study since all explants formed shoots. Up to 50% of the shoots rooted when cultured in 1/2 strength MS medium supplemented with 4.0mg.l^-1 IBA (Table 1). Rooting of E. caesia and E. macrocarpa micro-shoots has been achieved using 1/4 strength MS medium (Hartney, 1983). The mean number of roots increased with IBA concentration, up to an optimum of 0.8mg.l^-1 IBA. Higher concentrations, however, also encouraged callus proliferation. Callusing increased with time in culture and it was found necessary to maintain the shoots in the rooting medium for only about 3 weeks to prevent shoots being overgrown by callus. In the tissue culture of Pinus nigra, shoots had to be transferred to a hormone-free medium after about 4 weeks to prevent them from being overgrown with callus tissue (Kolevska-Pletikapic et al., 1983). Root initiation in NAA treatments was sporadic, though roots were observed as early as the second week (Table 2). Ahuja (1983) demonstrated the development of roots in in vitro cultured Aspen in Aspen Culture Medium (ACM) lacking in m-inositol and lysine, but supplemented with low levels of auxin such as NAA (0.1mg.l^-1) and IBA (0.5mg.l^-1). Indole acetic acid was a poor root initiator and only a low percentage of the shoots rooted when cultured in medium with 2.0mg.l^-1 IAA (Table 3). Roots were often feeble and cultures overgrown by callus. Some plantlets were successfully transferred into a 3:1 sterile mixture of sand and top soil. Further to the information presented here, other culture conditions likely to affect differentiation of shoots from juvenile propagules of mature Eucalyptus grandis such as light, media formulations, etc., may require further investigation.The next step would be to define optimal culture conditions for reliably rooting the in vitro propagated shoots and hardening them for field experimentation. ACKNOWLEDGEMENT The author wishes to thank technical staff of the Botany Department of the Tea Research Foundation of Kenya for technical help. This paper is presented with permission from the Director, Tea Research Foundation of Kenya. REFERENCES Ahuja, M.R. 1983. Somatic cell differentiation and rapid clonal propagation of Aspen. Silvae Genetica 32:131-135 Bonga, J.M. 1987. Clonal propagation of mature trees: Problems and possible solutions. In: Cell and Tissue Culture in Forestry. Vol.1. General Principles and Biotechnology. Bonga, J.M. and Durzan, D.J. (Eds.) pp. 249-271. Martinus Nijhoff publishers, Dordrecht, Netherlands. Boulay, M. and Franclet, A. 1983. Micropropagation de clones ages d'Eucalyptus selectionnes pour leur resistance au froid.In:Colloq. Int. Sur. Les. Eucalyptus resistance au Froid. IURFO-CSIRO 26-30 September 1983, pp 587-601. Burger, D.W. 1987. In vitro micropropagation of E. sideroxylon. Horticultural Science 22 : 496-497. Chaperon, H. 1975. Amelioration genetique des Eucalyptus hybrides au Congo-Brazaville. In: Third World Consultation on Forest Tree Breeding. Canberra, Documents (FO-FTD-77-4/9). Creswell, R.J. and Nitsch, C. 1975. Organ culture of Eucalyptus grandis L. Planta 125: 87-90. Curir, P., Damiano, C., Volpi, L. and Ruffoni, B. 1987. Observations on the development of new shoots from axillary buds in the in vitro propagation of Eucalyptus gunnii Hook. Horticultural Abstracts 57:929. Damiano, C., Curir, P. and Cosmi T. 1987. Short note on the effect of sugar on the growth in vitro of Eucalyptus gunnii. Horticultural Abstracts 57:919. Doorenbos, J. 1965. Juvenile and adult phases in woody plants. In: Encyclopedia of Plant Physiology XV/1. Ruhland, W. (Ed.), pp 1222-1235. Springer, Berlin. Durand-Cresswell, R., Boulay, M. and Franclet, A. 1982. Vegetative propagation of Eucalyptus. In: Tissue Culture in Forestry. Bonga, J.M. and Durzan, D.J. (Eds.), pp. 150-180. Martinus Nijhoff, The Hague. Durzan, D.J. and Campbell, R.A. 1974. Prospects for the introduction of traits in forest trees by cell and tissue culture. New Zealand Journal of Forestry Science 4:261-266. Durzan, D.J. and Chalupa, V. 1974. Growth and metabolism of cells and tissue of Jack pine (Pinus banksiana). Growth of cells in liquid suspension cultures in light and darkness. Canadian Journal of Botany 54:456-467. Eldridge, K.G. 1976. Breeding systems, variation and genetic improvement of tropical eucalypts. In: Tropical Trees: Variation, Breeding and Conservation. Burley, J. and Styles, B.T.(Eds.). Academic Press, London, Linnean Society Symposium series No.2. Erickson. T. 1965. Studies on the growth measurements of cell cultures of Haplopappus gracilis. Physilogia Plantarum 18:976-993. FAO. 1981. Eucalypts for planting. Food and Agriculture Organisation Forestry Series No. 11 pp. 204, Rome, Italy. Fossard, R.A. De, Nitsch, C., Cresswell, R.J. and Lee, E.C.M. 1974. Tissue and organ culture of Eucalyptus. New Zealand Journal of Forestry Science 4:267-278. Gupta, P.K., Mascarenhas, A.F. and Jagannathan, V.1981. Tissue culture of forest trees: clonal propagation of mature trees of Eucalyptus citriodora Hook, by tissue culture. Plant Science Letters 20:195-201. Hartney, V.J. 1982. Tissue culture of Eucalypts. Combined Proceedings of the International Plant Propagators Society 32:98-109. Hodgson, L.M. 1974. The breeding of Eucalypts in South Africa. Journal of the South African Forestry Association 89:13-15. Hodgson, L.M. 1976a. Some aspects of flowering and reproductive behaviour in Eucalyptus grandis (Hill) Maiden. 1. Flowering, controlled pollination methods, pollination and receptivity. South African Forestry Journal 97:18-23. Hodgson, L.M. 1976b. Some aspects of flowering and reproductive behaviour in Eucalyptus grandis (Hill) Maiden. 2. The fruit, seed, seedlings, self-fertility, percent selfing and inbreeding effects. South African Forestry Journal 98:32-33. Hsiao, K.C. and Bornman, C.H. 1989. Cyanide-initiated oxygen consumption in autoclaved culture medium containing sugars. Plant Cell Reports 8:90-92. Jalil, K. and Gavinlertvatana, P. 1989. Scale-up tissue culture system for mass propagation of mature Eucalyptus camaldulensis. Abstracts of a Seminar on Forest Tree Tissue culture, 17th-18th August 1989, National University of Singapore, Singapore. Kikuta, Y. and Okazawa, Y. 1984. Control of root and shoot bud formation from potato tuber cultured in vitro. Physiologia Plantarum 61:8-12. Kolevska-Pletikapic, B., Jelaska, S. and Berljak, J. 1983. Bud and shoot formation in juvenile tissue culture of Pinus nigra. Silvae Genetica 32:115-119. Konar, R.N. and Nagmani, R. 1974. Tissue culture as a method for vegetative propagation of forest trees. New Zealand Journal of Forestry Science 4:279-290. Kozai, T. 1991. Micropropagation under photoautotrophic conditions. In: Micro-propagation : Technology and Application. Debergh, P.C. and Zimmerman, R.H. (Eds.), pp. 447-469. Kluwer Academic Publishers, Dordrecht. Marino, G., Bertazza, G., Magnanini, E. and Altan, A.D. 1993. Comparative effects of sorbitol and sucrose as main carbon energy sources in micropropagation of apricot. Plant Cell Tissue and Organ Culture 34:235-244. Martin, S.M. and Rose, D. 1976. Growth of plant cells (ipomea) suspension cultures at controlled pH levels. Canadian Journal of Botany 54:1264-1270. Mascarenhas, A.F., Hazara, S., Potdar, U., Kulkarni, D.K. and Gupta, P.K. 1982. Rapid clonal propagation of mature forest trees through tissue culture. In: Plant Tissue Culture. Fujiwara, A. (Ed.), pp. 719-720. Proceedings of the 5th International Congress of Plant Tissue and Cell Culture. Tokyo. Matsumoto,T., Okunishi, K. Nishida, K. and Noguchi, M. 1972. Effects of physical factors and antibiotics on the growth of higher plant cells in suspension culture. Agricultural Biological Chemistry Journal 36:2177-2183. McComb, J.A. and Bennet, I.J. 1986. Eucalypt (Eucalyptus spp.) In:Biotechnology in Agriculture and Forestry Vol. 1.Trees. Bajaj Y.P.S. (Ed.), pp. 340. Springer Verlag, Berlin, Heidelberg. Minocha, S.C. 1987. pH of the medium and growth and metabolism of cells in culture. In: Cell and Tissue culture in Forestry. Vol. 1. General Principles and Biotechnology. Bonga, J.M. and Durzan, D.J. (Eds.), pp 125-144. Martinus Nijhoff Publishers, Dordrecht, Netherlands. Moncousin, C., Ribaux, M., Orke, J. and Gavillet, S. 1992. Effects of type of carbohydrate during proliferation and rooting of microcuttings of Malus Jork 9. Agronomie 12:775-781. Mozalewski, R.L. and Hackett, W.P. 1979. Cutting propagation of Eucalyptus ficifolia using cyto-kinin-induced basal trunk sprouts. Proceedings of the International Plant Propagators Society 29:118-124. Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 14:473-497. Nadgir, A.L., Phadka, C.H., Gupta, P.K. Parsharami, V.A., Nair, S. and Mascarenhas, A.F. 1984. Rapid multiplication of Bamboo by tissue culture. Silvae Genetica 38:219-223. Oka, S. and Ohyama, K. 1982. Sugar utilization in mulberry (Morus alba L.) bud culture. In:Plant Tissue Culture. Fujiwara, A. (Ed.), pp. 67-68. Proceedings of the 5th International Congress of Plant Tissue and Cell Culture, Tokyo. Oka, S. and Ohyama, K. 1986. Mulberry (Morus alba L.). In:Biotechnology in Agriculture and Forestry Trees. pp. 384-392 Springer-Verlag, Berlin. Paton, D.M., Willing, R.R., Nicholls, W. and Pryor, L.D. 1970. Rooting of stem cuttings of Eucalyptus:A rooting inhibitor in adult tissue. Australian Journal of Botany 18:175-183. Pua, E.C. and Chong, C. 1984. Requirement for sorbitol (D-glucitol) as carbon source for in vitro propagation of Malus robusta No 5. Canadian Journal of Botany 62:1545-1549. Robbins, W.R. 1957. Gibberellic acid and reversal of adult Hedera to a juvenile state. American Journal of Botany 44:743-746. Romano, A., Noronha, C. and Martins-Loucao, M.A. 1995. Role of carbohydrates in micropropagation of cork Oak. Plant Cell Tissue and Organ Culture 40:159-167. Rouland, H. 1973. The effect of cyclophysis and topophysis on the rooting ability of Norway spruce cuttings. Forest Tree Improvement 5:21-41. Street, H.E. 1969. Knowledge gained by culture of organs and tissue explants. In:Plant Physiology: A Treatise. Vol VB. Steward, F.C. (Ed.), pp. 3-224. Academic Press, New York. Thompson, M. and Thorpe, T. 1987. Metabolic and non-metabolic roles of carbohydrates. In:Cell and Tissue Culture in Forestry. Vol 1. General Principals and Biotechnology. Bonga J.M. and Durzan, D.J. (Eds.), pp. 89-112. Martinus Nijhoff Publishers, Dordrecht. Thorpe, T.A. 1982. Carbohydrate utilization and metabolism. In: Cell and Tissue Culture in Forestry, Vol.1. General Principals and Biotechnology. Bonga, J.M. and Durzan, D.J. (Eds.), pp. 325-368. Martinus Nijhoff Publishers, Dodrecht. Thorpe, T.A. and Patel, K.R. 1984. Clonal Propagation: Advertitious buds. In:Cell Culture and Somatic Cell Genetics of Plants. Vol.1. Vasil, I.K. (Ed.), pp. 49-60. Academic Press, New York. Von Arnold, S. and Eriksson, T. 1981. In vitro studies of adventitious shoot formation in Pinus contorta. Canadian Journal of Botany 59:870-874. Wainwright, H. and Scrase, J. 1989. Influence of in vitro pre-conditioning with carbohydrates during the rooting of microcuttings on in vitro establishment. Scientia Horticulturae 38:261-267. Welander, M., Welander, N.T. and Brackman, A.S. 1989. Regulation of in vitro shoot multiplication in Syringa, Alnus and Malus by different carbon sources. Journal of Horticultural Science 64:361-366. Wetherell, D.F. and Dougal, D.K. 1976. Sources of nitrogen supporting growth and embryo-genesis in cultured wild carrot tissue. Physiologia Plantarum 37:97-103. Winton, L.L. 1970. Shoot and tree production from aspen tissue cultures. American Journal of Botany 57:904-909. Wyk, G. Van. 1977. Pollen handling, controlled pollination and grafting of Eucalyptus grandis. South African Forestry Journal 101:47-53. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97030g.gif] [cs97030a.gif] [cs97030j.gif] [cs97030b.gif] [cs97030e.gif] [cs97030i.gif] [cs97030d.gif] [cs97030c.gif] [cs97030f.gif] [cs97030h.gif] |
| |||||||||

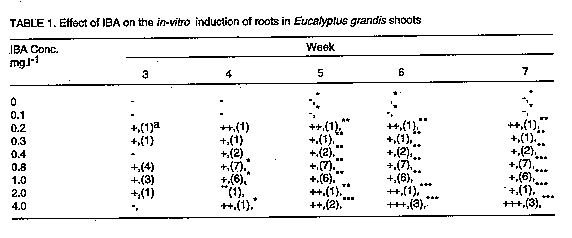{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}