 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Comparative response of a day-neutral and photoperiod-sensitive sorghum to delayed sowing or transplanting
A.Tenkouano, J. Chantereau^1, P. Sereme^2 and A.B. Toure^3 West and Central Africa Sorghum Research Network, International Crops Research Institute for the Semi-Arid Tropics, BP 320, Bamako, Mali. Present Address: International Institute of Tropical Agriculture, Oyo Road, PMB 5320, Ibadan, Nigeria c/o Lambourn & Co., Carolyn House, 26 Dingwall Road, Croydon, CR9 3EE, England
^1 Centre de Cooperation Internationale en Recherche Agronomique pour le
Developpment, International Crops Research Institute for the Semi-Arid
Tropics, BP 320, Bamako, Mali (Received 11 March, 1997; accepted 30 July, 1997)
Code Number: CS97032
Sizes of Files:
Text:
Graphics: Tables and line drawings (gif) - 43.1K
ABSTRACT
Soil moisture deficit frequently occurs at the onset of the growing season and may cause crop failure. Resowing with early maturing varieties has often been the only alternative, besides transplanting. To assess the relative efficiency of the two rescue methods two sorghum (Sorghum bicolor [L.] Moench) cultivars, 'IRAT204' (day-neutral) and 'Nazongala' (photoperiod-sensitive), were grown in seven drought-prone environments in Western Africa. Grain yield of both cultivars was reduced by delayed sowing or transplanting at all environments. However, transplanting had less depressive effects than late sowing for both cultivars. Yield reduction was lowest when transplanting or resowing was done not later than two weeks from normal sowing time. Compensation levels were higher with the cultivar 'IRAT204' than with 'Nazongala'. While transplanting could contribute to partial yield loss recovery under erratic rainfall environments, it should not be considered as an option beyond four weeks from the normal sowing dates with day-neutral cultivars, due to their lack of phenological plasticity. Key Words: Erratic rainfall, photoperiodism, yield loss compensation, Sorghum bicolor, transplanting RESUME Les deficits hydriques interviennent frequemment en debut d'hivernage et causent souvent des manques a la levee. En dehors du repiquage, le seul recours des paysans est de resemer avec des varietes plus precoces. En vue de comparer l'efficacite relative des deux methodes de sauvetage, deux varietes de sorgho (Sorghum bicolor [L.] Moench), 'IRAT204' (non photosensible) et 'Nazongala' (photosensible), ont ete evaluees dans sept sites en Afrique de l'Ouest. Les rendements obtenus avec le resemis ou le repiquage ont ete certes moins eleves que ceux resultant du semis a la date optimale, mais le niveau de compensation a ete plus important avec le repiquage, particulierement lorsqu'il avait ete fait au plus tard deux semaines apres la date normale de semis. Les taux de compensation partielle de rendement ont ete plus eleves chez 'IRAT204' que chez 'Nazongala'. Bien que le repiquage puisse contribuer a une reduction des pertes en zones de pluviometrie erratique, il est deconseille au-dela de quatre semaines apres la date normale de semis, notamment pour les varietes non photosensibles, du fait de leur manque de plasticite phenologique. Mots Cles: Pluviosite erratique, compensation de perte de rendement, Sorghum bicolor, repiquage INTRODUCTION
Drought frequently causes deficits in food crop production, resulting in undernutrition and starvation in several sub-Saharan countries (Johnson et al., 1992). The probability of drought in these countries is highest at the onset of rains and at the end of the cropping season (ICRISAT, 1984). Moisture deficits early in the season result in delayed and/or poor plant stand establishment (Soman et al., 1984). Terminal drought may cause severe yield loss, particularly when it occurs while plants are at the flowering stage (Grant et al., 1989; NeSmith and Ritchie, 1992).
Drought management through genetic and agronomic improvement is a major concern of crop research in many sub-Saharan countries. When soil moisture deficit occurs primarily during a certain stage of growth, genetic improvement may be for stage-specific drought tolerance. In many places, however, deficits can occur at anytime during the crop season and genetic improvement may be through either pyramiding stage-specific drought tolerance genes or selection of traits that improve stress avoidance or tolerance throughout the season.
Short season varieties may be used for late sowing (Catherinet et al., 1973). An alternative to sowing of short season varieties is the use of varieties which are phenologically plastic due to photoperiod sensitivity (Acosta-Gallegos and White, 1995). Resowing may not be an efficient option due to both climatic constraints and lack of adequate reserves of sowing seeds of good quality (Tenkouano and Sereme, 1996). Transplanting seedlings from a small and easily managed nursery is another option.
The objectives of this study were to determine the effects of late sowing and transplanting on the performance of phenologically plastic and inplastic sorghum cultivars under conditions of variable moisture availability. MATERIALS AND METHODS
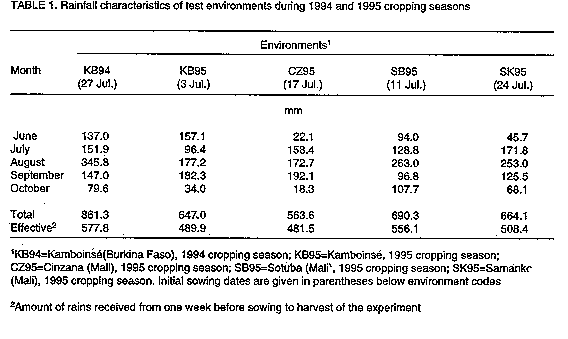
This study was conducted in a total of seven rainfed environments at four locations: Kamboinse, Burkina Faso (1 degree 32'W, 12 degrees 16'N), with and without tied ridges, in 1994 and 1995; and in three locations in Mali including Cinzana (3 degrees 56'W, 13 degrees 18'N), Samanko (8 degrees 7'W, 12 degrees 33'N),and Sotuba (7 degrees 56'W, 12 degrees 39'N). (Table 1). Treatments and experimental design. Two widely adapted sorghum varieties, IRAT204 and Nazongala (Tenkouano, 1995) were used in the trials. IRAT204 is an early maturing, short statured, daylength neutral caudatum variety. Nazongala is a medium maturity, tall, photoperiod-sensitive guineense landrace. The performance of these varieties was evaluated across seven planting methods. The control consisted of sowing at the onset of the rainy season and additional sowings or transplanting were delayed by 14, 21, and 28 days. Transplanted seedlings were from a nursery which was set up at the time of the first sowing. Thus, transplanted seedlings were 0, 14, 21, and 28 days older than field germinated seedlings from the corresponding sowing times.
The experimental design was a split-plot arranged within a randomised complete block design with three replications. Main plot and subplot treatments consisted of the planting methods and cultivars, respectively. Experimental units were three rows with lengths of 5 m in Burkina Faso, and 6 m in Mali. Sowing or transplanting was done at a spacing of 75 cm between rows and 20 cm within rows in Burkina Faso. Spacing was 80cm between rows and 30 cm within rows in Mali. In both countries, preplanting fertilisation (15N:23P2O5:15K2O) was done at the rate of 100 kg ha^-1 and urea (46N:0P2O5:0K2O) was applied as side-dressing in two split-doses of 50 kg ha^-1 each approximately 30 and 60 days after planting. Weed control was done mechanically as needed.
Data collection and statistical analysis. Data were recorded on number of days from planting to flowering, plant height to the flag leaf, grain yield, average number of seeds per plant, and 1000 seed weight. For transplanting treatments, days to flowering were calculated as the number of days from transplanting to flowering plus the number of days in the nursery.
A combined analysis of variance was carried out using the GLM procedure in SAS (SAS, 1985). Both main and subplot effects were considered fixed within random environments and replications. Statistical tests of significance were done using the procedures of Nevado and Ortiz (1985). When interactions effects were significant, the data were subjected to principal component analysis (PCA) using an additive main effects and multiplicative interaction (AMMI) model (Gauch, 1992). RESULTS AND DISCUSSION
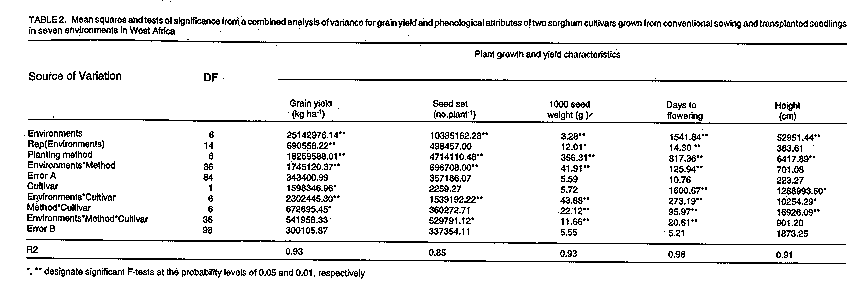
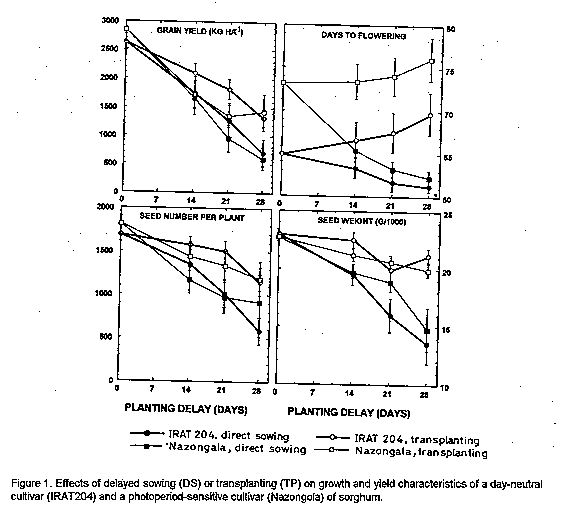
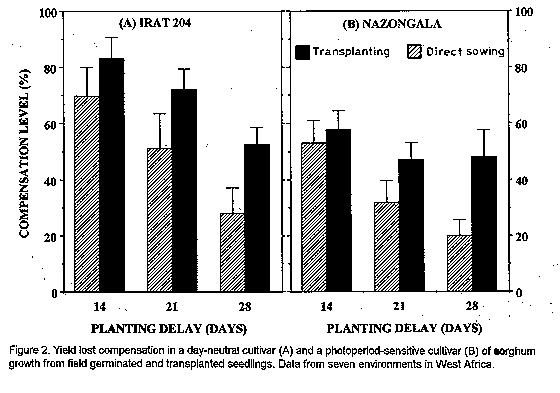
Crop environment and planting method significantly (P <0.05) affected all traits, but cultivar effects were not significant for seed number and kernel weight (Table 2). Two-factor or three-factor interaction effects were significant for most traits, except plant height and seed number. Plant height was not significantly affected by the interaction of environments and planting methods or by the interaction between these factors and cultivars (Table 2). Seed number was most affected by environment, followed by planting method and environment x cultivar effects (Table 2). Grain yield of both cultivars was reduced by delayed sowing and transplanting in all environments, but yield reduction was less with transplanting, due to both higher seed set and higher seed weight, than with direct sowing (Fig.1). Compensatory growth was best when sowing or transplanting was delayed by only 14 days: with sowing, compensation levels were 70 % and 52 % for IRAT204 and Nazongala, respectively, whereas transplanting resulted in 82 % and 57 % compensation for these cultivars, respectively. Compensation levels were also higher with IRAT204 than with Nazongala with 21 day delay, regardless of the planting method, and with sowing with 28 day delay, but compensatory growth from 28-day-old transplanted seedlings was equivalent in both cultivars (Fig. 2). Delayed sowing reduced the time to flowering in both cultivars,while transplanting resulted in delayed flowering in both cultivars (Fig.1). For both cultivars, transplanted seedlings took more time to reach the flowering stage than seedlings from direct sowing, despite the fact that transplanted seedlings were older than their field germinated counterparts. This was attributed to transplanting shock, a phenomenon reported in other studies (Berner et al., 1996). This may be responsible for the reduced compensatory growth expressed by plants derived from late transplanting.
Genotypic differences in response to photoperiod may cause ontogenic differences in sorghum (Quinby, 1973) and other crop species (Major, 1980). The ontogeny of sorghum consists of three developmental phases: emergence to floral inititiation (GS1) which determines time to flowering, floral initiation to anthesis (GS2) which determines the number of seeds, and anthesis to physiological maturity (GS3) which determines seed weight (Eastin, 1972). In this study, the number of seeds per plant was reduced by delayed sowing, particularly in the photoperiod-sensitive cultivar (Fig. 1), probably due to shortening of the GS2 stage. Delayed transplanting also reduced the number of seeds in both cultivars. However, reduction in the photoperiod-sensitive cultivar was less severe when transplanting was delayed by 28 days, which was also associated with greater yield recovery (Fig. 1). This suggests that the transplanting shock observed with younger seedlings may be due to interruption of the process of seed number determination in the GS2 phase.
Seed weight was little affected by transplanting in both IRAT204 and Nazongala. However, delayed sowing significantly reduced seed weight, particularly in IRAT204 (Fig.1). Due to its lack of phenological plasticity, the GS3 of the photo-period-insensitive cultivar coincided with periods of heavy midge (Contarinia sorghicola Coq.) infestation in all environments, which adversely affected both seed set and grain filling.
Grain yield was significantly affected by two-factor and three-factor interaction effects. Interaction between environments and cultivars or planting methods was attributed to differences in weather conditions and planting time (Table 1), and differences in cultivar response to photoperiod. Late planting resulted in lower performance of the photoperiod- sensitive cultivar, due to smaller seed number resulting from reduced development time. With early planting (e.g. Kamboinse 1995), the photoperiod-sensitive cultivar had the highest yield. Another major component of the interaction was midge infestation which affected late plantings of both cultivars. However, effects were more pronounced on the photoperiod- insensitive plot-1 at Kamboinse in 1994.
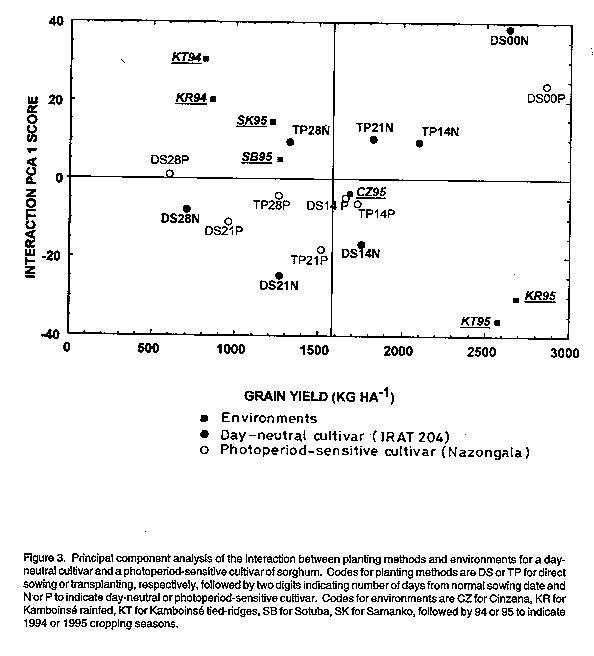
PCA analysis of the interactions between planting methods and environments indicated that deviation from the expected grain yield was lowest with transplanting 14 days from normal sowing time for both cultivars (Fig. 3). Also, direct sowing with 14 day delay was equivalent to transplanting with 21 day delay. However, crop performance was less predictable with direct sowing for the day-neutral cultivar than for the photoperiod-sensitive cultivar as evidenced by the large IPCA scores (Fig. 3). Beyond 21 days from normal sowing time, transplanting provided a more predictable crop performance than direct sowing for both cultivars, even when sowing was done one week earlier than transplanting (Fig. 3). Thus, transplanting is a better option than direct sowing within three weeks of the normal planting date. However, transplanting should not be considered as an option beyond four weeks from the normal sowing dates with the photoperiod- insensitive cultivar since it expressed less than half of its potential yield. In contrast, compensation level was essentially the same for the photoperiod-sensitive cultivar, regardless of the delay period (Fig. 2), which may be indicative of its phenological plasticity.
Phenological plasticity is often thought to be the rationale for the selection and cultivation of photoperiod-sensitive cultivars under subsistence farming conditions (Curtis, 1967; Dogget, 1988; Zongo and Tenkouano, 1995). However, grain yield reduction was more severe in the photoperiod-sensitive cultivar compared to the day-neutral cultivar due to reduced seed number. Is it possible that photoperiodism has simply evolved as a mechanism for self-perpetuation rather than productivity per se?
Regardless of the physiological mechanisms of phenological plasticity, this study also suggests that, compared with delayed direct sowing, the cultivar with zero yield recorded at all Malian locations in 1995 and yields of less than 100 g use of transplanted seedlings allows for reduction of hierarchical relationships which could develop among plants near the time of emergence (Volenec et al., 1987; Counce et al., 1989) and cause high heterogeneity in growth and maturity.
We conclude that transplanting, preferably within 14 days but not later than 21 days from normal sowing time, is a feasible option that could contribute to partial yield loss recovery under erratic rainfall environments, particularly with photoperiod-insensitive cultivars. Clearly, therefore, transplanting could provide a more reliable management option than late sowing for crop production under risk-avoidance subsistence farming systems.
REFERENCES Acosta-Gallegos, J.A. and White, J.W. 1995. Phenological plasticity as an adaptation by common bean to rainfed environments. Crop Science 35:199-204. Berner, D.K., Kling, J.G. and Singh, B.B. 1996. Striga research and control - A perspective from Africa. IITA Research 13:15-21. Catherinet, M.D., Dumont, S. and Mayaki, A.A. 1973. Le mil et le sorgho dans l'agriculture du Niger. A. Besoins, sol et climat, facteurs limitants. L'Agronomie Tropicale 18:108-113. Counce, P.A., Moldenhauer, K.A.K. and Marx, D.B. 1989. Rice yield and plant yield variability responses to equidistant spacing. Crop Scence 29:175-179. Curtis, D.L. 1967. The races of sorghum in Nigeria: Their distribution and relative importance. Experimental Agriculture 3:275-286. Dogget, H. 1988. Sorghum. Longman Scientific and Technical, New York. 512pp. Eastin, J.D. 1972. Photosynthesis and translocation in relation to plant development. In: Sorghum in the Seventies. Rao, N.G.P. and House, L.R. (Eds.), pp. 214-246. Oxford and IBH publishing Co. New Delhi, India. Grant, R.F., Jackson, B.S., Kiniry, J.R. and Arkin, G.F. 1989. Water deficit timing effects on yield components in maize. Agronomy Journal 81:61-65. Gauch, H.G. Jr. 1992. Statistical Analysis of Regional Yield Trials: AMMI Analysis of Factorial Designs. Elsevier. Amsterdam, The Netherlands. 289pp. ICRISAT. 1984. Agrometeorology of Sorghum and Millet in the Semi-Arid Tropics. Proceedings of an International Symposium, 15-20 Nov 1982, ICRISAT Center, India. Patancheru, India. 322pp. Johnson, J.J., Allredge, J.R., Ullrich, S.E. and Dangi, O. 1992. Replacement of replications with additional locations for grain sorghum cultivar evaluation. Crop Science 32:43-46. Major, D.J. 1980. Photoperiod response characteristics controlling flowering of nine crop species. Canadian Journal of Plant Science 60:778-784. NeSmith, D.S. and Ritchie, J.T. 1992. Effects of soil water-deficits during tassel emergence on development and yield components of maize (Zea mays L.). Field Crops Research 28:251-256. Nevado, M. and Ortiz, R. 1985. Prueba de hipotesis en serie de ensayos. Agro-Ciencia 1:23-37. Quinby, J.R. 1973. The genetic control of flowering and growth in sorghum. Advances in Agronomy 25:125-162. SAS Institute. 1985. SAS Users' guide: Statistics. Version 5 Edition. SAS Institute Inc., Cary, NC, U.S.A. Soman, P., Peacock, J.M. and Bidinger, F.R. 1984. A field technique to screen seedling emergence in pearl millet and sorghum through soil crust. Experimental Agriculture 20:327-334. Tenkouano, A. 1995. West and Central Africa Sorghum Research Network Annual Report-1994.ICRISAT, Bamako, Mali.54pp. Tenkouano, A. and Sereme, P. 1996. Some implications of genotype x environment interactions on sowing seed quality of sorghum. African Crop Science Journal 4: 11-18. Volenec, J.J., Cherney, J.H. and Johnson, K.D. 1987. Yield components, plant morphology, and forage quality of alfalfa as influenced by plant population. Crop Science 27: 321-326. Zongo, J.D. and Tenkouano, A. 1995. Etude de la diversite chez les sorghos du Burkina Faso. Processing and Industrial Utilization of Sorghum and Related Cereals in Africa. Proc. Regional Symposium, 22-26 Nov. 1993, Ouagadougou, Burkina Faso. In: Menyonga, J.M., Bezuneh, T., Nwasike, C.C., Sedego, P.M. and Tenkouano, A. (Eds.), pp. 237-245. OAU/STRC-SAFGRAD. Ouagadougou, Burkina Faso.
Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97032e.gif] [cs97032b.gif] [cs97032a.gif] [cs97032c.gif] [cs97032d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}