 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Physiological response of groundnut to plant population density J.P. AKWILIN TARIMO Department of Crop Science and Production, Sokoine University of Agriculture, P. O. Box 3005, Morogoro, Tanzania (Received 25 September, 1996; accepted 12 August, 1997)
Code Number: CS97033
Sizes of Files:
Text:
Graphics: Tables and line drawings (gif) - 19.5K
ABSTRACT
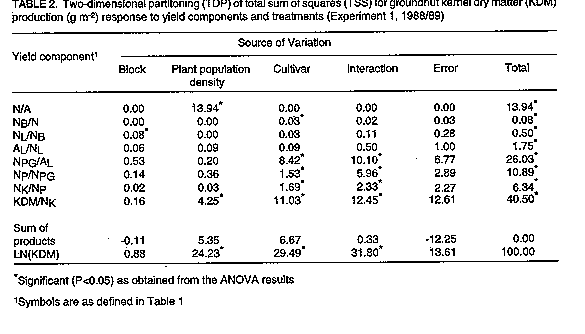
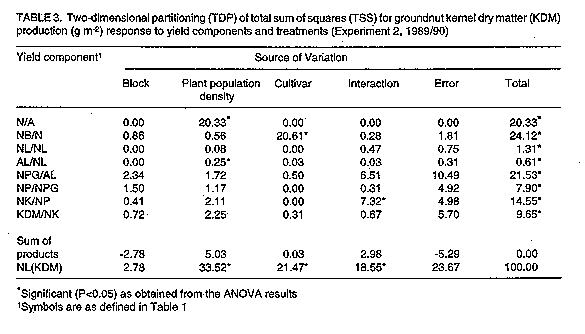
Two experiments were carried out in two seasons at the University of Queensland, Redland Bay Farm in Southeast Queensland, Australia to identify some physiological determinants of groundnut response to plant population density after anthesis. The first experiment was grown in square spacings while the second one was grown in rectangular spacings. Yield componet analysis (YCA) techniques were used to study the yield and yield component responses to the different levels of plant population density. The results show that variation in kernel yield m^-2 among the plant population densities was associated with responses of some reproductive yield components, e.g. kernel mass (KDM/Nk) to the plant population densities. In contrast, the responses of vegetative components to those densities had little association with the overall kernel yield differences in this study. It is concluded that the two-dimensional partitioning (TDP) technique of YCA provides an effective method of identifying physiological and morphological determinants of yield variation in groundnut under field conditions. From the results, variation in kernel dry matter production was associated with responses of KDM/NK, Npg/Al, NPG, NK/NP and NB/N to the treatments. Key Words: Arachis hypogaea L., lernel dry matter, plant population density, two-dimensional partitioning, yield components RESUME Deux experimentations ont ete menees pendant deux campagnes a l'Universitede Queensland, Redland Bay Farm au Sud-est de Queensland, en Australie afin d'identifier quelques determinants physiologiques de la reaction de l'arachide a la densite de la population des plantes apres l'anthse. La premire experience etait menee aux ecartements carres tandis que la deuxime etait menee aux ecartements rectangulaires. Les techniques d'analyse des composants de rendement (YCA) etaitent utilisees pour etudier le rendement et la reaction des composants de rendement en fonction de niveaux differents de la densitede la population des plantes. Les resultats ont demontre que la variation en rendement des graines m^-2 parmi les densites de la population des plantes etait associee aux reactions de quelques composants reproductifs de rendement e.g. le rapport masse des graines (KDM/Nk) - densite de la population des plantes. Par contre, les reactions des composants vegetatifs ces densites-la avaient des effets minimes en rapport avec les differences en rendement des graines dans leur ensemble. On peut conclure que la technique de la division a double dimension (TDP) de la technique de (YCA) pourvoit une methode efficace pour identifier des determinants physiologiques et morphologiques de la variation de rendement d'arachide en conditions de champ. De ces resultats, on peut deduire que la variation dans la production des matieres seches des graines etait liee la reaction des traitements de KDM/NK, Npg/A1; NPG, NK/NP et NB/N. Mots Cles: Arachis hypogaea L., matires seches des graines, densite de la population des plantes, division a double dimension, composants de rendement INTRODUCTION Groundnut (Arachis hypogaea L.) is cultivated widely in the arid and semi-arid regions of the world (40 degrees North and 40 degrees South) in temperature regimes ranging from warm temperate to equatorial. It is an important oilseed of the semi-arid tropics. Groundnut yields in the semi-arid tropics are low (0.5 to 1.0 ton ha^-1), compared with 2.7 tons ha^-1 in the United States of America (U.S.A). Future improvement of groundnut yields world-wide might depend on detailed analyses of the genetic potential of the current cultivars at varying plant population density in order to ascertain sustenance of yields under farm conditions (Donald, 1963; Byth et al.,1987). The genetic potential of crop plants can be regorously studied by applying yield component analysis (YCA) procedures (Eaton et al., 1986). Besides plant population density, yield components of groundnut are also affected by environmental factors such as soil physical and chemical properties, temperature and the available soil moisture (Ishag, 1985). However, little information is available on the physiological determinants of groundnut response to plant population density. The most frequently studied yield components of the crop are pod and kernel numbers and mass (Bell et al., 1987). Vegetative yield components such as branches and leaves have only recently been analysed in relation to plant population density response of crops. This paper reports on the interaction of vegetative and reproductive components in determining yield variation in groundnut grown at different plant population densities. The main objective was to contribute to an understanding of the physiological response of groundnut to agronomic practices. MATERIAL AND METHODS Two experiments were carried out in two seasons, 1988/89 and 1989/90 at the University of Queensland's Redland Bay Farm (27 degrees 37' S; 153 degrees 17' E) in Australia. The area has a deep, friable, fertile red loam soil (Krasnozem) with 60% clay, 15% silt and 25% sand (Keating, 1981). Six groundnut cultivars belonging to two varieties of the cultivated groundnut were studied. The cultivars were Improved Virginia Bunch (IVB), NC7 and Q18801 (Virginia types) and TMV-2, McCubbin and Red Spanish (Spanish types). In both seasons, experiments were established on land previously under a fallow crop of sorghum (Sorghum bicolor L. Moench). The land was ploughed one month before planting. Five days before planting, a compound fertiliser was broad-cast at the rate of 25 kg N, 30 kg P and 25 kg K ha^-1. Both experiments were planted by hand in November of each season to match the cropping season of East Queensland. The spacings for the first experiment were 0.1 x 0.1, 0.2 x 0.2, 0.3 x 0.3 and 0.4 x 0.4 m. These spacings produced populations of 100, 25, 11.1 and 6.3 plants m^-2, respectively. The experiment was laid out in split-plots in a randomised complete block design (RCBD) with 3 replication. The plot sizes were 4.0 x 1.3, 4.0 x 2.3, 4.0 x 3.9 and 4.0 x 5.2 m, from the highest to the lowest plant population density, respectively. For the second experiment, each cultivars was sown in rows 0.6 m apart and four within-row spacings of 0.04, 0.06, 0.08 and 0.16 m which resulted in populations of 42, 28, 21 and 10 plants m^-2, respectively. The layout of the experiment was factorial in a RCBD with three replications. The plot size was constant for all plant population densities, i.e., 3.0 x 3.0 m. Plots were maintained weed-free throughout the season using a combination of mechanical and hand weeding operations. A broad spectrum insecticide, Lannate (225 gl^-1 methomyl) was sprayed at the recommended rate when foliar pests were observed on the crop. Attempts were made to ensure that Lannate was used only when there was an evidence of insect pest damage to avoid environmental contamination. Foliar diseases were controlled by frequent application of Bravo (500 gl^-1 Chlorothalonil) at the rate of 2 ml l^-1 of water, particularly during periods of high humidity and following heavy rainfall. Apart from rust (Puccinia arachidis) which occurred towards the end of the season, foliar disease were not important in both years. Six harvest were carried out in Experiment 1, and four in Experiment 2. At each harvest, six adjacent plants in a row were harvested from each plot (Yayock, 1979), taking care to ensure that there was no loss of pods. The data recorded included leaf area (AL), number of branches (NB), leaves (NL), pegs (NPG), pods (NP) and kernels (NK) per plant. Other data included dry mass (DM) of stems (WST), leaves (WL), pods (WP) and kernels (WK) per plant. Plant parts were used as yield components to study variation in yield among the groundnut cultivars. A yield component in this study was defined as the ratio of one plant part to another or to the whole plant. Thus, the following ratios were used as yield componets: number of plants (N) per area (A) (N/A); number of branches per plant (NB/N); leaf number per branch (NLNB); area per leaf (AL/NL); peg number per leaf area (NPG/AL); ratios of pod number to peg + pod number (NP/NPG); kernel number to pod number (NK/NP); and, individual kernel dry mass (KDM/NK). These yield components were selected and arranged in a chronological sequence to determine their associations with KDM m-2 development in groundnut. Thus kernel dry matter production was considered to be a mathematical product of the following ratios: N/A x NB/N x NL/NB x AL/NL x NPG/AL x NP/NPG x NK/NP x KDM/NK = KDM (g m^-2) In this model, it was assumed that KDM production would depended on the plant population densities (N/A), branches per plant (NB/N), leaves per branch (NL/NB), area per leaf (AL/NL), peg + pod number per leaf area (NPG/AL), pegs per peg+pod number (NP/NPG), kernels per pod (NK/NP) and kernel (KDM/NK). The ratio, NPG/AL, provides a link between assimilate source and assimilate sinks during reproductive growth, while N/A converts data on a per plant basis to per unit area basis. Symbols used are those proposed by Jolliffe et al. (1982). The geometric model above was transformed into an additive model by taking natural logarithms of the ratios (Eaton et al., 1986). The additive model was then analysed by stepwise multiple regression procedures and the two-dimensional partitioning (TDP) technique. In both experiments, pre-flowering data were excluded from the model to avoid inclusion of a zero-yield component in the sequence. Statistical analyses of the data were carried out using either SAS (SAS Institute Inc.) programme or the University of Queensland, Department of Agriculture Statistical package, MicroQUASP, on an IBM-compatible microcomputer. RESULTS The data summarised in all tables in this paper were computed as ratios of total sum of squares (SS) for orthogonal components to total SS for KDM (yield), from the results of analysis of variance (ANOVA). These rations were then multiplied by 100 to obtain the coefficients of determination (%) (Eaton et al., 1986). The significance levels indicated were based on the ANOVA results. Table 1 shows the relationships between KDM and the yield components as were influenced by the plant population densities and cultivars at three consecutive harvests after anthesis in groundnut for Experiment 1 (square arrangement). The results show that variation in kernel yield at early pod growth [91 days after planting (DAP)] was accounted for mainly by the variation in KDM/NK (66%) and NK/NP (19%). Later in reproductive growth (113 DAP), the variation in kernel yield was associated with variations in N/A (22%) and KDM/NK (60%). At kernel maturity, yield differences were determined by KDM/NK (41%), NPG/AL (26%), N/A (14%), NK/NP (6%) and NP/NPG (11%). Overall, vegetative yield components accounted for less of the variation in kernel yield than reproductive components (Table 1). Low variation in vegetative components and greater variation in reproductive components promoted kernel yield differences among treatments (Table 1). In the second experiment (rectangular arrangement), variation in kernel yield at early reproductive growth (83 DAP) was caused by the variation in NB/N while at kernel maturity, N/A (20%) and NB/N were important yield components. This was in addition to other components, including NL/NB (24%), NP/NPG (22%), NK/NP (14%) and KDM/NK (10%) which also significantly affected KDM at maturity (Table 1). The results of TDP analysis for Experiment 1 (Table 2) show clearly that the total sum of squares (TSS) for kernel yield at maturity were mainly accounted for by the differences in plant population densities (24%), cultivars (29%) and their interaction (32%). The main components in this case were N/A (14%) and KDM/NK (4%) (Table 2). The other components were less affected by the plant population density treatments. Kernel yield differences among cultivars were associated with the variation in KDM/NK (11%) and NPG/AL (8%). The other components were similar in all cultivars. In the second experiment, plant population density accounted for 34% and cultivars 21% of the total variation in kernel yield at maturity (Table 3). Treatment interaction accounted for 19% of that variation. The contribution by plant population density to the variation in kernel yield was mainly through the variation in N/A (20%) and AL/NL (0.3%). No other component was significantly affected by the plant population density in this experiment (Table 3). The contribution by cultivars to the variation in yield was associated with variation in NB/N (21%). As observed in Experiment 1, previously, increased number of branches per plant decreased kernel yield (Table 3). These results suggest that factors promoting vegetative growth after anthesis in groundnut, such as low plant population density, would greatly reduce kernel yield at maturity.
DISCUSSION The objective of a crop improvement programme in agriculture is to maximise productivity at maturity under field conditions. In many crop plants, YCA has provided key indicators for determining sources of physiological and genetic variations in yield (Genter and Camper, 1973; Geadelman and Peterson, 1976; Cross, 1977; Eaton, et al., 1986; Kuriakose and Joseph, 1986). In the two experiments, variation in kernel yield during reproductive growth was accounted for by the plant population densities. At maturity, however, the importance of plant population density (N/A) on kernel yield declined. This gave way to reproductive components to express greater roles in determining yield variation among the groundnut cultivars. Significant components of kernel yield in the groundnut are NPG/AL, NP/NPG, NK/NP, KDM/NK and NB/N (Table 1). These components were considered useful as genetic determinants of variation in kernel yield among the cultivars. When treatments and yield components of kernel yield were analysed by the TDP technique, the results showed that, in both seasons, the plant population densities accounted for over 26% of the variation in kernel yield, mainly through N/A and KDM/NK (Tables 2 and 3). Differences among cultivars accounted for an average of over 25% of the variation in kernel yield in both seasons, mainly through KDM/NK and NB/N. Treatment interactions were responsible for over 26% of that variation in KDM at maturity. The main components of variation in this case were NPG/AL, KDM/NK, NP/NPG and NK/NP (Tables 2 and 3). The most significant contribution of this study has been in underlining the usefulness of the TDP technique of yield component analysis (Eaton et al., 1986) in tracing sources of variation in kernel yield among the groundnut cultivars grown under different plant population densities. While TDP is yet to be fully applied by the majority of plant breeders and plant physiologists in identifying traits useful for crop yield improvement, there is a clear indication that the method simplifies the overall process of studying plant allometric relationships under different cultural practices. CONCLUSION The yield component analysis techniques used in this study effectively identified components associated with groundnut response to different plant population densities under field conditions. Thus, the following yield components were associated with yield differences observed among the varieties in the different plant population densities; KDM/NK, NPG/AL, NP/NPG, NK/NP and NB/N, in that order. These are considered as useful physiological and morphological determinants of groundnut response to agronomic practices under farm conditions. ACKNOWLEDGEMENT This paper is part of a Ph.D. thesis submitted to the University of Queensland, Australia, under the supervision of Dr. F.P.C. Blamey. The research was funded by the Australian International Development Assistance Bureau (AIDAB). REFERENCES Bell, M.J., Muchow, R.C. and Wislon, G.L. 1987. The effect of plant population in peanuts (Arachis hypogaea L.) in a monsoonal tropical environment. Field Crops Research 17:91-107. Byth, D.E., Shorter, R. and Surmarno, 1987. Genetic limits to improvement of food legumes. In: Food Legume Improvement for Asian Farming Systems: Proceedings of an International Workshop at Khon Kaen. Thailand 1-5 September 1986. Wallis, E.S. and Byth, D.E. (Eds.). ACUAR Proceedings 18:156-168. Cross, H.Z. 1977. Interrelationships among yield components in early maize. Crop Science 17: 741-745. Donald, C.M. 1963. Competition among crop and pasture plants. Advances in Agronomy 15:1-18. Eaton, G.W., Bowen, P.A. and Jolliffe, P.A. 1986. Two-dimensional partitioning of yield variation. Horticultural Science 21:403-408. Jolliffe, P.A., Eaton, G.W. and Lovett Doust, J. 1982. Sequential analysis of plant growth. The New Phytologist 92:287-296. Keating, B.A. 1981. Environmental effects on growth and development of cassava (Manihot esculenta Crantz) with special reference to photoperiod and temperature. Ph.D. Thesis, University of Queensland. Kuriakose, P.K. and Joseph, C.A. 1986b. Path analysis and selection index in groundnut. Agricultural Research Journal of Kerala 24: 101-110. Yayock, J.Y. 1979. Effects of variety and spacing on growth, development and dry matter distribution in groundnut (Arachis hypogaea L.) at two locations in Nigeria. Experimental Agriculture 15:339-351. Copyright 1997 The African Crop Science Society The following images related to this document are available:Line drawing images[cs97033c.gif] [cs97033b.gif] [cs97033a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}