 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
IMPROVING THE EFFICIENCY IN BREEDING COWPEA FOR ADAPTATION IN SEMI-ARID TROPICS N. MULEBA, J.T. OUEDRAOGO^1, I. DRABO^1 and C. DABIRE^1
IITA-SAFGRAD, 01 B.P. 1495 or 1783, Ouagadougou 01, Burkina Faso (Received 4 June, 1995; accepted 19 September, 1997)
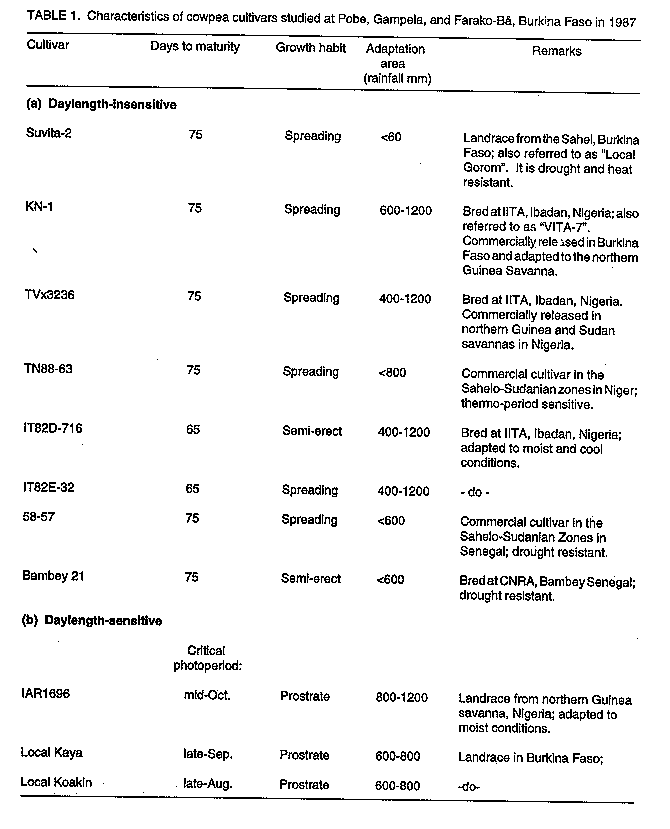
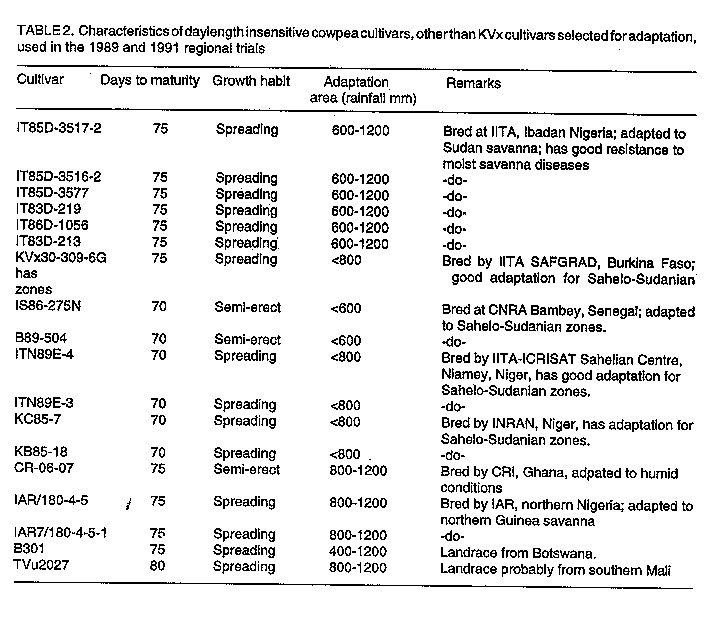
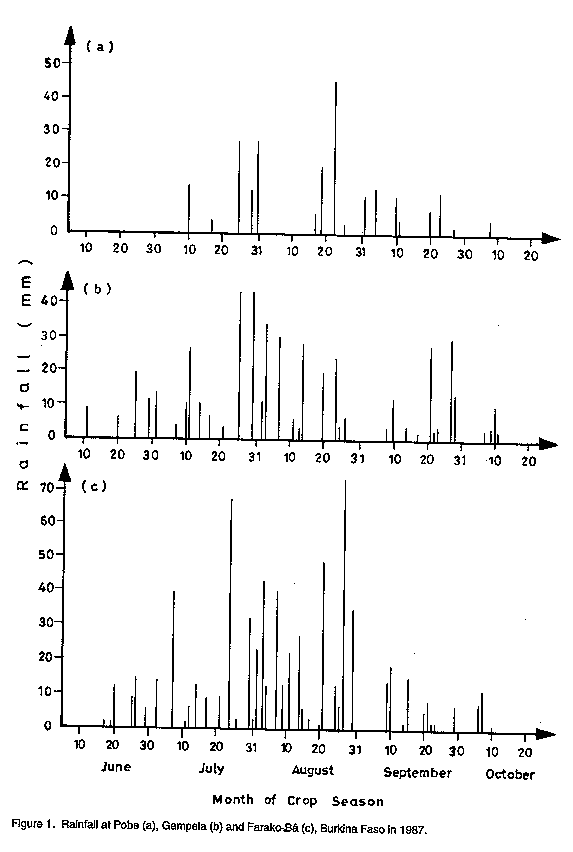
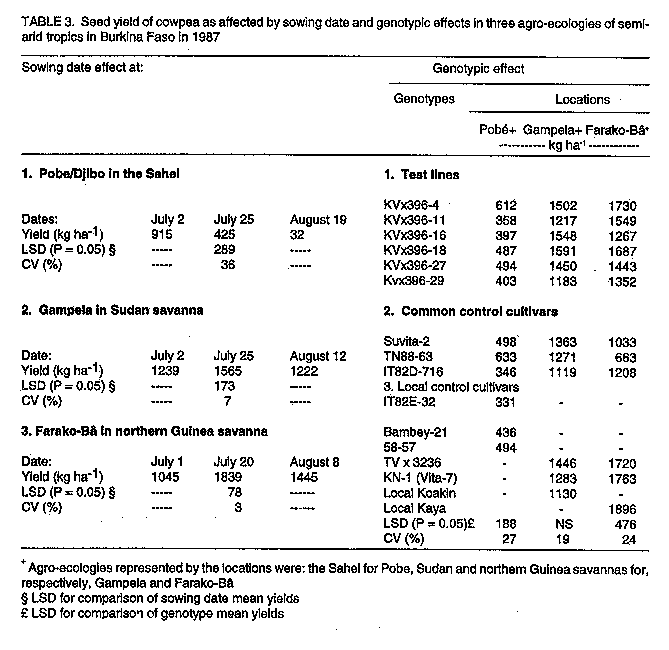
Code Number:CS97039 ABSTRACT Crops in semi-arid tropics (SAT) are subject to yield losses due to drought and heat stress as well as excess moisture. Cultivars in these ecologies should be genetically buffered against environmental variations to ensure food security. A strategy for combining, in a cross, genes for high yield, tolerance to drought, heat and excess moisture, and resistance to associated diseases is presented. The research objective was to improve, over currently used breeding approaches, the efficiency in developing high yielding and highly adapted cowpea (Vigna unguiculata L. Walp.) cultivars. To this end, crosses were made involving parents of various characteristics and origins. Offsprings from the crosses were first screened for disease resistance. They were then subjected to a successional sowing cultivar evaluation (SSCE) for the identification of productive and adapted genotypes and were further evaluated for adaptation in various SAT in Burkina Faso. Selected recombinant genotypes were further evaluated for adaptation in various SAT ecologies in West and Central Africa. New, higher yielding and better adapted genotypes were identified only from crosses that involved high yielding parents adapted to extreme conditions in the different ecologies in the sub region. Some of the recovered genotypes showed stable, high yields and good adaptation in specific and also across different ecologies in semi-arid West and Central Africa. This indicated that the breeding strategy used was effective for the development of cultivars highly adapted to semi-arid tropical conditions. Key Words: Adaptation, breeding, drought, excess moisture, resistance, semi-arid tropics, stability, tolerance, Vigna unguiculata, yield RESUME La culture du niebe (Vigna unguiculata L. Walp) en zones semi-arids tropicales subit d'importantes chutes de rendements subsequantes a la secheresse et/ou au stress thermique ainsi qu'a l'exces d'humidite. Les cultivars diffuses dans ces zones ecologiques doivent etre genetiquement tamponnes contre les aleas climatiques afin d'y securiser la production alimentaire. Une strategie: d'apport, par croisements, d'un ensemble des genes fonctionnels pour la haute productivite et la resistance/tolerance a la secheresse, au stress thermique et a l'exces d'humidite y compris les maladies qui y sont associees en vue de favoriser, par brassage, la formation des genotypes de recombinaison tres productive et a adaptation tres large est ici presentee. L'enjeu de cette recherche etait d'ameliorer l'efficacite du developpement de nouveaux cultivars de niebe hautement productifs et bien adaptes. A cet egard, des croisements impliquant des parents des caracteristiques et d'origines diverses ont ete effectues. Les descendants issus de ces croisements ont d'abord ete cribles pour la resistance aux maladies. Ensuite, ils ont ete evalues au Burkina Faso en utilisant differentes dates de semis. Ceci afin de les exposer aux conditions variables de l'environment et de selectionner ceux ayant un bon comportement tout en etant hautement productifs. Les genotypes ainsi selectionnes ont en-outre fait l'objet d'evaluation dans diverses ecologies de la zone semi-aride de l'Afrique du Centre et de l'Ouest. Quelques genotypes de recombinaison, hautement productifs et bien adaptes, ont ete recouvres seulement dans les populations issues des croisements dont les parents etaient tres productifs et adaptes aux conditions extremes dans les differentes ecologies de la zone semi-aride. Certains de ces genotypes ont exhibe non seulement un haut rendement stable, mais aussi une bonne adaptation tant aux conditions specifiques qu'a travers les differentes ecologies semi-arides en Afrique du Centre et der L'Ouest. II apparait donc que la strategie utilisee a ete tres efficace pour le developpement de nouveaux cultivars de niebe a haute productivite et bien adpates aux conditions semi-arides tropicales. Mots Cles: Adaptation, amelioration, exces d'humidite, rendement, resistance, secheresse, stabilite, tolerance, Vigna unguiculata, zones semi-arides tropicales INTRODUCTION Semi-arid tropics (SAT) in West and Central Africa have been experiencing climatic change since the 1970s characterised by erratic, below average and poorly distributed rainfall (Turk et al., 1980; Virmani et al., 1980; Tucker et al., 1991). Severe cowpea yield losses may result from moisture shortage, high temperatures, or both under drought conditions (Turk et al., 1980; Warrag and Hall, 1983, 1984). Muleba et al. (1991) showed that dry spells in West and Central Africa were associated with both high ambient air and soil temperatures (measured at a 5 cm depth). Soil temperatures greater than 38 C hindered cowpea growth, development, and productivity (Lal, 1975; Muleba, 1988; Muleba et al., 1991). Excess moisture during rainy years may also have a detrimental effect on cowpea yields due to soil moisture saturation, disease outbreaks, or both (Muleba et al., 1991). Therefore, commercial cultivars in SAT should be genetically buffered against environmental variations to mitigate yield losses and ensure food production security for farmers. This can be achieved only if such cultivars possess genes for tolerance to both deficient and excess moisture conditions. Breeding cowpeas for adaptation in SAT has been difficult. Results obtained have been inconsistent and no progress has been made with regard to drought tolerance or resistance (Leleji, 1988). Similar difficulties are also encountered with other crops under semi-arid conditions (Zavala-Garcia et al., 1992; Calhoun et al., 1994). These difficulties may be attributed to either or both of the following: (i) the lack of recombinant genotypes in offspring which carry genes controlling high yield and tolerance to drought and heat as well as excess moisture; and (ii) large genotype x environment interaction variances that must be dissociated from genotypic variance in order to properly assess cultivars for high yield and adaptation. Zavala-Garcia et al. (1992) and Calhoun et al . (1994) proposed exposing genotypes to contrasting environments for a proper assessment of their genotypic value for yield and adaptation under semi-arid conditions. The genotype x environment interaction was successfully separated from the genotypic variances in SAT in West and Central Africa by Aggarwal and Haley (1988). He used a yield stability analysis based on regional cultivar evaluation trials conducted at eight locations in four countries over a two-year period. He was able to differentiate cultivars adapted to specific ecologies in the subregion. The cost and tedium of evaluating genotype x environment interaction and genotypic variances in multilocation trials can be reduced as proposed by Muleba et al. (1991). They used successional sowing, where each sowing date simulates a different set of variable environmental conditions including moisture level and air and soil temperatures to identify cultivars adapted to the Sudan Savanna ecology of SAT in West Africa with only a single year's data. Widely adapted recombinant genotypes can thus be efficiently recovered using this approach as compared to the current breeding methods, which are based on evaluation of offspring in trials and frequently conducted, for early generations, at one location using a single, optimal sowing date. Since a comprehensive approach for breeding cowpea for high yield and adaptation to SAT has not been tackled, we developed a strategy of combining, in a cross, genes for high yield and tolerance to drought, heat, excess moisture and resistance to associated diseases. To achieve this, we crossed three parents each tolerant to either drought and heat or excess moisture. Other crosses, involving parents of various adaptation characteristics, were also made. After screening offspring for field resistance to prevailing diseases, we subjected them to a single plant selection for high yield and wide adaptation using a successional sowing cultivar evaluation (SSCE) in early or late generations. We evaluated the selected promising lines against currently available adapted cultivars including the commercial ones used in different ecologies in Burkina Faso and other West and Central African countries. The purpose of this research was to improve, over current cowpea breeding approaches practised in the sub region, the efficiency for developing new, high yielding cultivars widely adapted to SAT conditions. This is vital for insuring food security to resource-poor peasant farmers. The progress achieved is discussed in this paper. MATERIALS AND METHODS Agroclimatic zones. In West and Central Africa, semi-arid tropics extend from 9o30'N to 15o00'N and from the Atlantic coast in the west to Eastern Chad in the east. They include three distinct ecologies: the Sahel, with rainfall of 200-600 mm from late June to early September, the Sudan Savanna, with 600-900 mm of rainfall from early June to late September; and the Northern Guinea Savanna, with rainfall of 900-1200 mm from late May to mid-October (Virmani et al., 1980). Predominant soils under rainfed agriculture are aridisols and alfisols in the Sahel and Sudan Savanna and alfisols, oxisols, and some aridisols in the Northern Guinea Savanna. Breeding and selection The KV x 396 cross. Three parents were used: (i) Suvita-2, a day length insensitive (DI), drought and heat resistant cultivar adapted to the Sahel but susceptible to diseases of the moist savanna (Muleba et al., 1991); (ii) KN-1, a high yielding DI cultivar, adapted to humid conditions, tolerant to excess moisture, resistant to disease of the moist savanna but highly susceptible to drought and heat stress; and (iii) IAR1696, a day length sensitive cultivar, adapted to the moist savanna zone with reactions to drought and excess moisture conditions similar to KN-1. They were subjected to a three-way cross, ((IARI696 x KN-1) x Suvita-2), in December 1982. The cross was named: "KV x 396." The objective of this cross was stated above in the introduction. Between mid-February to late July 1983 at Kamboinse, Burkina Faso, F1 and F2 seeds were sequentially grown in one and four 0.75 m^2 wooden boxes, respectively. The boxes were 20 cm deep, filled with soil to which animal manure (20% of the soil medium) wad added. They were kept in the greenhouse courtyard and exposed to direct sunshine in the ambient atmosphere. They were watered daily as needed and plants were protected against insect pests with insecticides. The F1 plants were harvested in bulk. Single F2 plants were harvested separately and the resulting F3 plants were each grown in a separate row in the Northern Guinea Savanna at Farako-Be, Burkina Faso from early August to mid-October 1983 (early August to mid-September is a protracted rainy period). Therefore, they were exposed to moist savanna fungal and bacterial diseases. Single, disease-free plants were harvested in rows (i.e., lines) that were less infected than others. Selected F4 lines were subjected to a single plant selection for resistance to cowpea mosaic virus disease. This was carried out in the Sudan Savanna zone at Loumbila, Burkina Faso during the off-season from mid-February to late May 1984 under irrigation. The disease, which is transmitted by aphids (Aphis craccivora Koch), can cause severe damage to cowpea in semi-arid West and Central Africa. Its incidence is very high at Loumbila during the off-season. Cultivar Suvita -2 is highly susceptible to this disease. Therefore, each line was grown in a single 2.0 m row plot bordered on each side by the parent Suvita-2, which served as a check and spreader row. The spacing was 0.75 m between rows and 0.2 m within a row. Single plants were selected only from the lines that exhibited good disease resistance as compared to Suvita-2. Selected F5 to F6 lines were subjected in 1984 to 1985, respectively, to single plant selection using the SSCE on the three dates each year: (i) late June, (ii) early to mid-July, and (iii) late July to early August. This was carried out at Pobe, which is 20 km south of Djibo, Burkina Faso in the Sahel. Only the 1985 experiment was replicated twice. The same field design was used as at Loumbila in 1984, except that the length of the single row plot, bordered by Suvita-2, was reduced to 1.0 m. Vigorous, high yielding, and disease free plants were separately selected only from lines that exhibited higher yield and better field disease resistance than the bordering Suvita-2 and other lines across sowing dates. A total of 10 F7 was selected at the end of the 1986 season and advanced to F8 through seed increase during the 1987 off-season. They were evaluated for yield and adaptation during the 1987 growing season against the best parent at two locations, Suvita-2 at Pobe in the Sahel and KN-1 at Kamboinse (16 km north of Ouagadougou) in the Sudan Savanna. A common control cultivar, TN88-63, was also used at both locations. The experimental design was the same as used in 1985-86, except that each genotype was grown in a three-row plot, 1.0 m long (Suvita-2 was thus eliminated in the bordering rows), and the trial was replicated twice at each location. Six advanced lines (KVx 396 lines no 4, 11, 16, 18, 27 and 29) were finally selected across dates and locations as the most productive and adapted lines. From this research, offspring from F3 to F8 were subjected to pedigree selection for their high genetic value in field disease resistance, high yield, and adaptation as compared to the population mean and the best parent. The six selected advanced lines were evaluated in 1987 at three locations, each representative of a specific ecological zone of the SAT in Burkina Faso. At each location, six cultivars were included as controls. They consisted of three controls common to all locations: Suvita-2 (best parent adapted to the Sahel only), TN88-63, and IT82D-716. Three controls specific for the ecological zone where the experiment was conducted were also used. They were: IT82E-32, Bambey-21, and 58-57 for the Sahel; TVx3236, KN-1, and local Koakin for the Sudan Savanna; and TVx3236, KN-1 (as the best parent), and local Kaya for the Northern Guinea Savanna. Cultivar IAR1696, the third parent, could not be tested because of its mid-October critical photo period. It requires regular precipitation throughout October for flowering, pod setting, and seed production. This cannot be guaranteed due to climatic change. Since the early 1970s, rainfall has tended to end in late September or early October. Similarly, cultivar KN-1 was not tested in the Sahel because of its high susceptibility to heat stress (Muleba, 1988; Muleba et al., 1991). The cultivars are described in Table 1. At Pobe in the Sahel, the experiment was conducted on a "typical" haplargid soil (deep to very deep silt, loam to clay loam). Seeds were sown at three dates: 2 and 25 July and 19 August 1987. In the Sudan Savanna, the experiment was conducted at Gampela, 20 km east of Ouagadougou, on an alfisol (sandy loam to sandy clay loam in the upper 45 cm layers of the soil). The sowing dates were 2 and 25 July and 12 August 1987. In the Northern Guinea Savanna, the experiment was sown at Farako-Be on a "rhodic-oxic" paleustalf soil on 1 and 20 July and 8 August 1987. The experiment was a split-plot design with four replications. Sowing dates were assigned as main-plots and cultivars as sub-plots. Each plot measured 4.5 x 5 m. Single superphosphate fertiliser at 21.8 kg P ha^-1 was broadcast on the field and incorporated into the soil before sowing. Seeds were sown two per hill and later thinned to one plant per hill two weeks after sowing. The spacing was 0.75 m between rows and 0.20 m between hills within a row. The crop was uniformly sprayed with the insecticides Monocrotophos (R) at 12 g a.i. ha^-1 when aphids (A. craccivora) were observed on the plants, Deltamethrin (R) at 12 g a.i.ha^-1 at flower bud formation, and a mixture of Deltamethrin (R) at 12 g a.i. ha^-1 and Endosulfan (R) at 400 g a.i. ha^-1 at 10 days after flowering for insect pest protection. Plots were kept free of weeds by hand-hoeing. Grain yields were assessed from the four central rows of each plot, leaving 0.50 m at the end of each row. Pods were harvested as they matured, air dried to a constant weight for 10 days, and threshed. Meteorological data. During the cropping season, rainfall was recorded daily at 8:00 a.m. near each experimental field from a standard rainfall gauge 1.5 m above ground level at the three locations in Burkina Faso. Other crosses. Thirteen other crosses were made KV x no164, 291, 293, 295, 305, 397, 398, 399, 401, 402 and 410. They involved cultivar B301 crossed to Suvita-2, TN88-63, 58-57, Tvx3236, some lines derived from Suvita-2, and also some Suvita-2-derived lines crossed to IT82D-716 and TVu2027. Offsprings of these crosses were first subjected to the classical breeding approach currently used in the sub region. This consisted of beginning selection with the F2 plant s and carrying it through the F8 generation at two locations, Kamboinse in the Sudan Savanna and Pobe in the Sahel. A single plant selection (pedigree selection) was made at each location for adaptation, good podding, seed type, and resistance to various field diseases. Selected advanced line (after F8), hitherto not subjected to evaluation for genetic value for high yield and adaptation, were screened at Kamboinse and Pobe in 1988 for adaptation using the SSCE. Three sowing dates were used at each location, the experiment was not replicated, and the highest yielding lines with good field disease resistance across sowing dates and locations were selected. The selected lines were evaluated in 1989 and 1990 for yield and adaptation in the three semi-arid ecologies represented in Burkina Faso. This included the above two ecologies used for selecting offspring and also the Northern Guinea Savanna at Farako-Be. The SSCE was used with three dates at each location, the experiment was replicated, and the best commercial cultivars were used as controls. Only the descendants of three crosses, KVx305 (KVx146-27-4 x KVx30-6246-2-5K), KVx398 (KVx61-74 x B301), and KVx402 (KVx30-166-36 x B301) were selected. These crosses shared the following in their parentage: genes for resistance to drought and heat from Suvita-2, and tolerance to excess moisture from TVx3236 and/or TVu2027, both of which are adapted to the Northern Guinea Savanna ecology. The best cultivar derived from KVxcrosses (no 396, 305, 398 and 402) were evaluated for yield and adaptation in regional trials conducted in SAT in West and Central Africa in 1989 and 1991-92. The 1989 regional trials. Two regional trials were conducted, for the Sahelo-Sudanian zone and the other for the Northern Guinea Savanna zone. The Sahelo-Sudanian trial involved the four best KVx396 cultivars (KVx396-4-4, KVx 396-4-5-2D, KVx396-18-10 and KVx396-11-6), selected as described above, and five other cultivars (KVx30-309-6G, IT85D-3517-2, IT85D-3516-2, IT85D-3577, and IT83D-219) described in Table 2, which are adapted to this ecological zone. They were tested against three controls. Two were common commercial cultivars (TN88-63 and TVx3236) also adapted to this ecological zone (Aggarwal, 1988; Muleba et al ., 1991), and one was a local commercial cultivar provided by each participating national programme. The trial was established at five locations: Kamboinse, Burkina Faso; Mouda, Cameroon; Gassi, Chad; Kolo, Niger; and Minjibir, Nigeria. The Northern Guinea Savanna trial included five KVx396 cultivars (KVx396-4-2, KVx 396-4-4, KVx396-4-5-2D, KVx 396-16-10, and KVx 396-18-10) and two other cultivars (IT86D-1056 and IT83D-213), which are adapted to this ecology and are described in Table 2. They were tested against two controls, KN-1, which is better adapted to this ecology, and a local commercial cultivar provided by each participating national programme. The trial was established at eight locations: Ndali/INA, Benin; Farako-Be, Burkina Faso; Sanguere, Cameroon; Bouake, Cote d'Ivoire; Yundum, the Gambia; Nyampkpala, Ghana; Zaria, Nigeria; and Tantiegou, Togo. The 1991 regional trials. Two other regional trials were conducted in 1991. The Sahelo-Sudanian trial, including the coastal savanna in Benin, involved the three best cultivars: KVx 396-4-5-2D (the outstanding entry in all ecologies of semi-arid in the 1989 regional trial), KV x 402-5-2, and KVx402-19-5. The latter two are advanced lines selected using the SSCE (described above for crosses other than KVx396). This trial also included the cultivar KVx164-41-64, an advanced line rejected for selection using the SSCE, and six other cultivars described in Table 2: B89-504N, IS86-275N, ITN89E-3, ITN98E-4, KC85-7, and KB85-18. These were bred for adaptation in the ecological zone by various national agricultural research systems (NARS). They were tested against two controls, TVx3236 and a local commercial cultivar. The trial was conducted at Ajouhoun/Porto Novo and Niaouli, Benin in the coastal savanna and in the Sahelo-Sudanian ecology at 11 locations: Kamboinse and Pobe, Burkina Faso; Gueringuew/Maroua and Mouda/Maroua, Cameroon; Douguis and Gassi, Chad; Cinzana and Koporo, Mali; Silla, Mauritania; Kolo, Niger; and Minjibir, Nigeria. The Northern Guinea Savanna trial also involved KVx396-4-5-2D, five cultivars (KVx305-2-118-23-2, KVx305-118-31, KVx398-7-1, KVx402-5-2, and KVx402-19-1) which were advanced lines selected using the SSCE, and three cultivars described in Table 2 and bred by various NARS for this ecological zone (CR-06-07, IAR7/180/-4-5, and IAR7/180-4-5-1). They were all tested against three controls: KN-1, TVx3236, and a local commercial cultivar. The trial was carried out at 13 locations in 1991: Farako-Be and Nyangoloko, Burkina Faso; Sanguere and Touboro, Cameroon; Poumbaidi/Paoua and Soumbe, Central African Republic; Ferke-ssedougou, Cote d'Ivoire; Somipa and Yundum, the Gambia; Katibougou, Mali; Samaru, Nigeria; and Ativeme and Tantiegou, Togo. In 1992, the trial was repeated at Zaria, Nigeria and Ativeme, Togo and conducted for the first time at Kankan, Guinea Conakry. Therefore, the 1991 trial was conducted under a total of 16 different conditions in the Northern Guinea Savanna. Each trial consisted of 12 entries in a 3 x 4 compact randomised complete block design replicated four times. The 1989 regional trial for the Northern Guinea Savanna was an exception. It consisted of a 3 x 3 compact randomised complete block, since it had only nine entries. The experimental plot had four rows, each 4 m long and 0.75 m apart. The trial was conducted upon request by the NARS. The agronomic practices used were those recommended for cowpea production in the area where the experiment was conducted. Cowpea pods were harvested at full maturity in the two central rows, air dried to a constant moisture content, threshed, and dry grains weighed. Statistical analysis Breeding and selection. Each trial was analysed separately by location using analysis of variance to test the significance of differences in sowing date, cultivar, and sowing date x cultivar interaction effects. When significant, the mean square for sowing date x cultivar (or genotype) interaction effect was used to test the significance of the genotypic effect as described by McIntosh (1983) for the split-plot design. Sowing dates were random and genotypes fixed. "Student" t-test were used accordingly with appropriate mean squares to establish significant levels. Regional trials. Regional trials were analysed separately by year for each location using analysis of variance to test the significance of cultivar effect. A combined analysis of variance across locations was also carried out to test the significance of differences in location, genotype, and location x genotype interaction effects (McIntosh, 1983). Locations were random and genotypes fixed. "Student" t-tests were used accordingly to establish significant levels. The Finally and Wilkinson (1963) model was applied to study the adaptation and yield stability of cultivars when the location x genotype interaction effect was significant. Regression analysis of the mean yield(s) of cultivars on condition indices was conducted. This allowed the computation of the slope (beta) and the coefficient of determination (r^2) associated with the regression line of the mean yields of cultivars. These were based on 5 and 13 conditions (i.e., 5 and 13 locations in 1989 and 1991, respectively, for the Sahelo-Sudanian trials) and 8 and 16 conditions (i.e., 8 and 13 + 3 locations in 1989 and 1991, respectively, including 1992 for the Northern Guinea Savanna trials). The r^2 measures the percentage of the sum of squares accounted for by the regression line. RESULTS Meteorological conditions. Rainfall data are reported for the 1987 line evaluation experiments at the following locations in Burkina Faso: Pobe in the Sahel, Gampela in the Sudan Savanna, and Farako-Be in the Northern Guinea Savanna (Fig. 1). A total of 225, 658 and 887 mm rainfall were received, respectively, at Pobe, Gampela, and Farako-Be. These corresponded to 56, 88, and 84% of the long-term (25 years) averages of 400, 750 and 1050 mm for each respective location (Virmani et al., 1980). Rainfall was poorly distributed at Pobe throughout the cropping season (Fig. 1 a). At Gampela, rainfall was scattered from late June through early July, in late July, and from late August through October (Fig. 1b). At Farako-Be, however, significant dry spells occurred mostly in September and October, while excess moisture conditions prevailed in August (Fig. 1c).
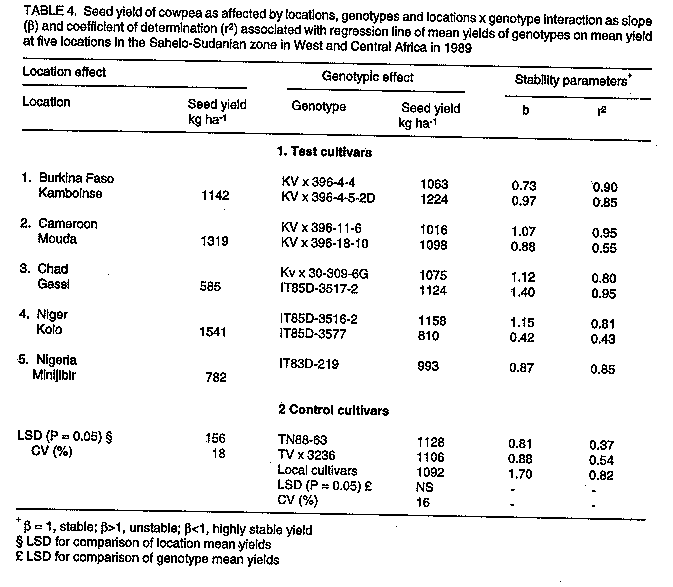
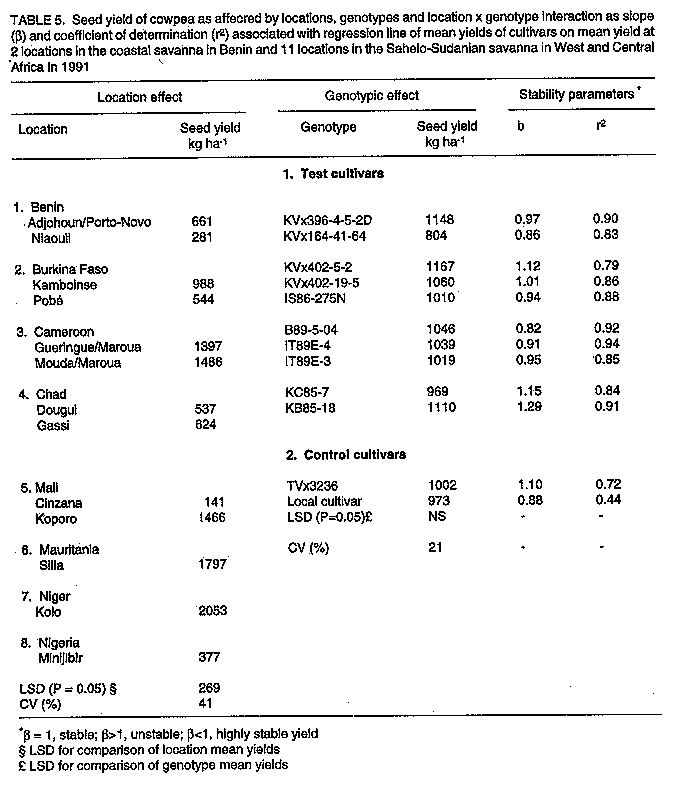
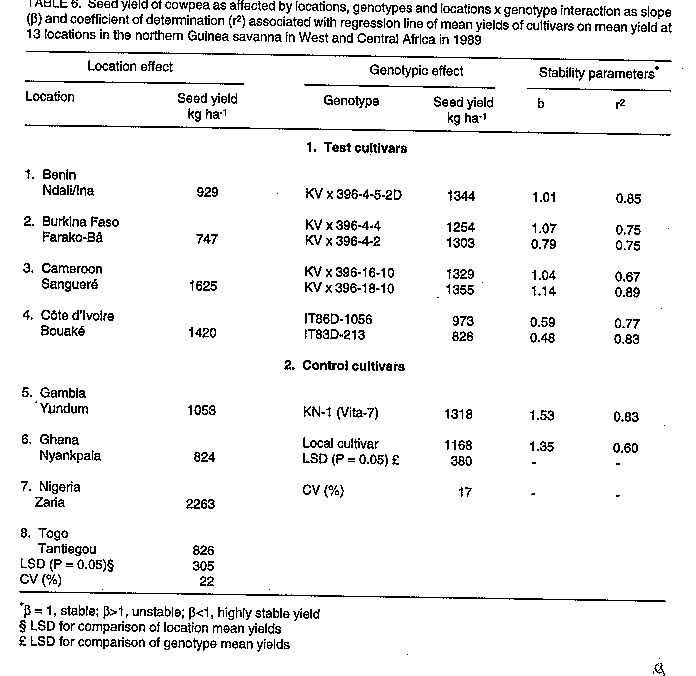
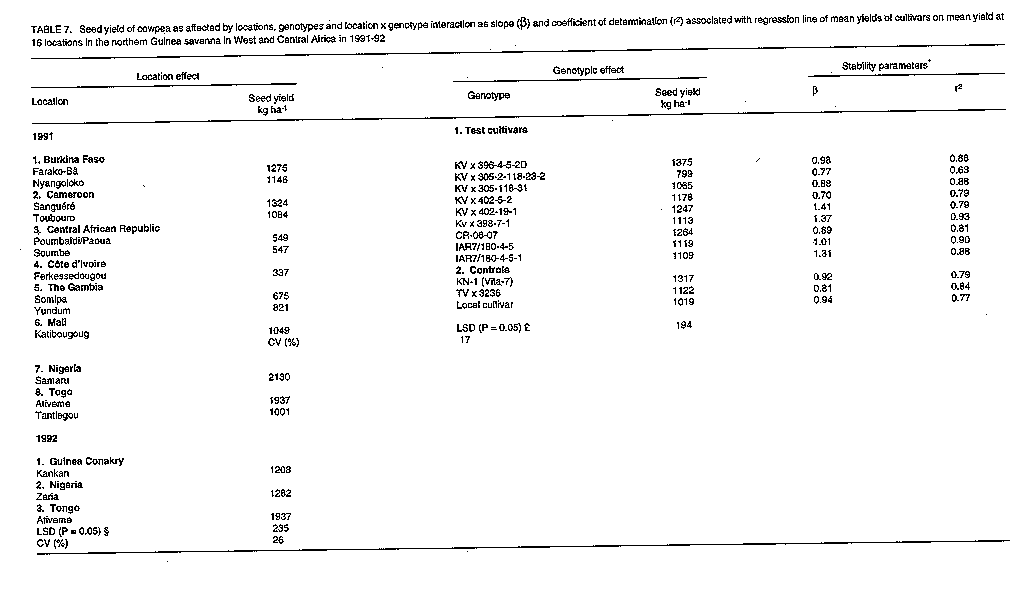
Breeding and selection. Sowing date, genotype, and sowing date x genotype interaction effects significantly affected cowpea yields in the three ecologies of SAT in Burkina Faso. The exception was the genotypic effect at Gampela. The effects of sowing dates and genotypes are given in Table 3. In the Sahel at Pobe/Djibo, dry spells in August and September (Fig. 1a) occurred during pod fill for the intermediate sowing date and during flowering, pod set and filling for the late sowing date (data not shown). This resulted in significant yield losses for these dates as compared to the first sowing date. End of season dry spells in September and October at Gampela and Farako-Be (Fig. 1b & c) caused significant yield losses for the first and late sowing dates as compared to the intermediate sowing date at both locations. The intermediate sowing date was also affected by the early September dry spells (Fig. 1b & c). However, since the dry period was before the initiation of flowering, the plants recovered quickly and suffered little damage. Lines KVx396-4 and KVx396-18 were the highest or among the highest yielding genotypes in the three ecologies. They contrasted with the three common control cultivars: Suvita-2, TN88-63, both of which exhibited good adaptation only in the Sahel and Sudan Savanna ecologies, and IT82D-716, which was not adapted at all to semi-arid conditions. Regional trials. In the Sahelo-Sudanian and coastal savanna ecologies, only the genotypic effect was insignificant (Table 4, Table 5). However, cultivars interacted differently with variable environmental conditions as shown by the slope (beta) and coefficient of determination (r^2). In the tables, beta = 1, beta > 1, and beta < 1 indicate stable, unstable and highly stable yields, respectively. The r^2 measured the percentage of the yield variations accounted for by the regression line, r^2 = 1.0 being 100%. Cultivar KVx164-41-64 together with IT85D-3577 and IT83D-219 in 1989 and KC85-7 and local control cultivars in 1991 were the lowest yielding cultivars (Tables 4, 5). Location, genotype, and genotype x location interaction effects were all significant in the Northern Guinea Savanna (Table 6, Table 7). The KVx396 cultivars and KVx 402-19-1 were either the highest or among the highest yielding cultivars. Similar results were obtained with cultivars KN-1 in 1989 and 1991-92, and CR-06-07 in 1991-92 only. These latter two cultivars were indeterminate. Therefore, they flowered and set and filled pods as long as it rained. They were different from all the KVx396's and KVx402-19-1, which behaved as determinate cultivars. This may explain the high yield and adaptation of KN-1 and CR-06-07 in the Northern Guinea Savanna ecology. DISCUSSION Insufficient rainfall and frequent dry spells associated with high soil temperatures, varying from 38 to 45 C and sometimes rising to 50 C (data not shown), prevented cultivars from fully expressing their yield potential in 1987 at Pobe, Burkina Faso (Table 3). Similar conditions, though less severe than in the Sahel, might have hindered cowpea productivity in early and late sowing at Gampela in the Sudan Savanna, and in early sowing at Farako-Be in the Northern Guinea Savanna during the same year. High soil temperatures (greater than 38 C, measured at a 5 cm depth) have been reported to have a detrimental effect on cowpea growth, development, and productivity even under no moisture stress (Lal, 1975; Muleba, 1988). When heat was combined with drought, only drought and heat resistant cultivars could survive and produce substantial yields (Muleba, 1988; Muleba et al., 1991). These severe conditions may explain the low yield in the Sahel, and to some extent, for the early and late sowing in the Sudan and the early sowing in the Northern Guinea Savannas of Burkina Faso in 1987. Lines KVx396-4, KVx396-27 and KVx 396-18 together with Suvita-2 (their best parent for the Sahel) and TN88-63, both drought and heat resistant (Muleba et al., 1991) controls, were the highest yielding lines and cultivars in the Sahel. This is evidence for the adaptation of these lines to drought and high temperature conditions. Excess moisture conditions in August 1987 at Farako-Be in the Northern Guinea Savanna (Fig. 1) resulted in an outbreak of two major diseases, brown blotch (Colletotrichum capsisi) and web blight (Rhizoctonia solani)). Cowpea mosaic virus, transmitted by aphids (A. craccivora), also infected cowpea plants at the same location. Cultivars Suvita-2, which is particularly susceptible to cowpea mosaic virus (Muleba et al., 1991), and TN88-63 both exhibited high susceptibility to all these diseases (data not shown). They gave the lowest yields in the Northern Guinea Savanna, in contrast with lines KVx396-4 and KVx396-18, which gave high yields (Table 3) and displayed good disease resistance. The high yields and wide adaptation expressed by lines KVx396-4 and KVx396-18 in all ecologies of SAT in Burkina Faso significantly differentiated them from the three common control cultivars, including Suvita-2. These results give evidence of the high genetic buffering capacity of these lines. They presumably inherited this genetic buffering capacity through recombination of favourable genes contributed by each parent in the cross. The successful genetic recombination of both lines is also well illustrated by the fact that these lines yielded equal to or better than the best parents (Suvita-2 in the Sahel and KN-1 in the Sudan and Northern Guinea Savannas) and the best control cultivars. Each of these cultivars was used in its ecology of adaptation. In regional trials, severe environmental conditions in the Sahelo-Sudanian and coastal savanna ecologies prevented cultivars from fully expressing their yield potential, as observed in the Sahel and Sudan Savanna in 1987 in Burkina Faso. However, the cultivars were differentiated in all ecologies of SAT in West and Central Africa by their ability to interact with variable environmental conditions. All KVx396 cultivars, perhaps with the exception of KVx396-18-10 in the Northern Guinea Savanna, exhibited stable to highly stable yields in all ecologies tested. The regression line of these cultivars accounted for most of the yield variations. They were highly adapted and interacted the least with variable environmental conditions. Other KVx cultivars (subjected to SSEC) ranged from unstable to highly stable yields. Their regression line also accounted for most of the yield variations. Those cultivars with stable to unstable yields were adapted under the ecological conditions in which they were tested, but unlike the KVx396 cultivars, they required favourable environmental conditions to produce high yields. In contrast, cultivars with highly stable yields: KVx164-41-64, KVx305-2-118-23-2, KVx305-118-31 and KVx402-5-2 (only in the Northern Guinea Savanna for the latter cultivar), also exhibited a trend toward low yields. Therefore, they were poorly adapted to the ecologies in which they were tested. Cultivars from the NARS and the International Institute of Tropical Agriculture (IITA) and the controls, except for IS86-275N, B89-504, IT89E-3, TVx3236 (Table 5), and KN-1 and CR-06-07 (Table 7), reacted to variable environmental conditions as did the KV x cultivars other than the KV x 396's. Among the poorly adapted cultivars, some required favourable environmental conditions to give relatively high yields, whereas the regression line of others did not account for most of the yield variations. These cultivars, like KVx cultivars other than the KVx396's, do not appear likely to guarantee food production security under unfavourable conditions. Cultivars IS86-274N and B89-509 from Senegal, IT89E-4 and IT89E-3, developed by the IITA for adaptation to Sahelo-Sudanian ecologies, and TVx3236, reported to be highly adapted to Sahelo-Sudanian ecologies (Aggarwal, 1988; Muleba et al., 1991), were as highly adapted as the KVx396 cultivars in Sahelo-Sudanian and the coastal savanna zones. Cultivars KN-1 and CR-06-07 in the Northern Guinea Savanna ecology yielded similar results. The high adaptation of the latter two cultivars in the Northern Guinea Savanna can be attributed to their indeterminate nature as stated earlier. Cultivars subjected to the SSCE either in early or late generations in the selection proved to be adapted particularly in the Sahelo-Sudanian and coastal savanna zones. This is in line with the resultant poor adaptation exhibited by cultivar KVx164-41-64 in the Sahelo-Sudanian and coastal savanna zones. Its poor performance was consistent with its rejection for selection for high yield and adaptation using the SSCE. The poor adaptation of KVx305, KVx398, and to some extent, KVx402 cultivars in the Northern Guinea Savanna could be attributed to the bulk of genes brought to the cross by their parents, TVx3236 and TVu2027, which are adapted to moist savanna conditions. Cultivar TVx3236, although reported to be adapted to SAT in West and Central Africa (Aggarwal, 1988; Muleba et al., 1991), was not consistently outstanding in the Northern Guinea Savanna ecology (Tables 3,6,7). In contrast TVu2027 is not known to be as high yielding as KN-1, TVx3236, or IAR1696 in the Northern Guinea Savanna ecology. Cultivar KVx396-4-5-2D produced high yields and expressed the highest adaptation in all ecologies in SAT in West and Central Africa. This showed its high yield and genetic buffering capacity against variable environmental conditions in SAT in Burkina Faso. The same was already exhibited in 1987 by line KV x 396-4, from which KVx396-4-5-2D was derived. Cultivars KVx 396-18-10 and KVx402-19-1 also produced high yields and expressed genetic buffering capacity in different ecologies in West and Central Africa, although they tended to require favourable environments for high yields. These results emphasise the need to include high yielding parents which are well-adapted to extreme environmental conditions in crosses for breeding for high yield and adaptation in SAT. Failure to achieve this would make breeding for adaptation to semi-arid conditions very difficult, as observed by Leleji (1988). This is also illustrated in this research by cultivar KVx164-41-64, and by all crosses other than KVx396, except to some extent, KVx402, KVx398. CONCLUSION The strategy of combining, in a cross, genes for high yield and high adaptation from parents highly adapted to extreme environmental conditions proved to be very effective. It promoted the formation of a recombinant genotype carrying favourable genes from each parent. The genotype was recovered during the selection process by exposing the offspring to contrasting environ-mental conditions during the same growing season using the SSCE. This was in agreement with the propositions by Zavala-Garcia et al. (1992) and Calhoun et al. (1994). In regional trials conducted in semi-arid West and Central Africa, the genotype was not only high yielding but also had a high genetic buffering capacity against variable environmental conditions. This is a guarantee for food security to resource-poor peasant farmers. This strategy, therefore, offers a new prospect in genetic manipulation for mitigating yield losses due to environmental hazards in SAT. ACKNOWLEDGEMENT The contribution of national scientists belonging to the SAFGRDA cowpea network from Benin, Burkina Faso, Cameroon, Central African Republic, Chad, Cote d'Ivoire, The Gambia, Ghana, Guinea Conakry, Mali, Mauritania, Niger, Nigeria and Togo is gratefully acknowledged. REFERENCES Aggarwal, V.D. and Haley, S.D. 1988. Breeding for drought and Striga resistance. In: Food Grain Production in Semi-arid Africa. Proceedings of an international Drought Symposium. Menyonga, J.M., Bezuneh, T. and Youdeowei, A. (Eds.), pp. 307-319. Trowbridge, UK. OAU/STRC-SAFGRAD (Redwood Brun Ltd). Calhoun, D.S., Gebeyehu, G., Miranda, A., Rajaram, S. and van Ginkel, M. 1994. Choosing evaluation environments to increase wheat grain yield under drought conditions. Crop Science 34:673-678. Finally, K.W. and Wilkinson, G.N. 1963. The analysis of adaptation in a plant breeding programme. Australian Journal of Agricultural Research 14:742-754. Lal, R. 1975. Role of mulching techniques in tropical soil and water management. IITA Technical Bulletin 1:1-38. Leleji, O.I. 1988. Cowpea breeding in Nigeria. In: State of Cowpea Research in Semi-arid zones of West and Central Africa. Proceedings of the first seminar for cowpea lead centre scientists; November 14-25, 1988; IITA-Ibadan, Nigeria. Muleba, N. and Emechebe, A.M. (Eds.), pp. 60-61. (IITA-SAFGRAD, OAU/STRC-SAFGRAD, Ouagadougou Burkina Faso). McIntosh, M.S. 1983. Analysis of combined experiments. Agronomy Journal 75:153-155. Muleba, N. 1988. Response of cowpea to high soil temperatures and drought. In: Food Grain Production in Semi-arid Africa. Proceedings of an international drought symposium. Menyonga, J.M., Bezuneh, T. and Youdeowei, A. (Eds.), pp. 331-349. Trowbrige, UK: OAU/STRC-SAFGRAD (Redwood Burn Ltd). Muleba, N., Mwanke, M. and Drabo, I. 1991. Use of successional sowing in evaluating cowpea (Vigna unguiculata) adaptation to drought in the Sudan savannah zone. 1. Seed yield response. Journal of Agricultural Science, Cambridge 116:73-81. Turk, J.K., Hall, A.E. and Asbell, C.M. 1980. Drought adaptation of cowpea. 1. Influence of drought on seed yield. Agronomy Journal 72:413-420. Tucker, C.J., Dregne, H.E. and Newcomb, W.W. 1991. Expansion and contraction of the Sahara desert from 1900-1990. Science 253:299-301. Virmani, S.M., Reddy, S.J. and Bose, M.N.S. 1980. Manuel de climatologie pluviale de I'Afrique Occidentale: Donnees pour des stations selectionees. ICRISAT Information Bulletin no 7, pp. 56. Warrag, M.O.A. and Hall, A.E. 1983. Repro-ductive responses of cowpea to heat stress: genotypes differences in tolerance to heat at flowering. Crop Science 23:1088-1092. Warrag, M.O.A. and Hall, A.E. 1984. Repro-ductive responses of cowpea (Vigna unguiculata (L.) Walp) to heat stress. II. Responses to night air temperatures. Field Crop Research 8:17-33. Zavala-Garcia, F., Bramel-Cox, P.J., Eastin, J.D., Witt, W.D. and Andrews, D.J. 1992. Increasing the efficiency of crop selection for unpredictable environments. Crop Science 32:51-57.
Copyright 1997, African Crop Science Society The following images related to this document are available:Line drawing images[cs97039a.gif] [cs97039e.gif] [cs97039c.gif] [cs97039h.gif] [cs97039d.gif] [cs97039f.gif] [cs97039b.gif] [cs97039g.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}