 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
SEASONAL FLUCTUATIONS AND DAMAGE OF LEPIDOPTERAN STEMBORERS OF MAIZE IN A MAJOR AGROECOZONE OF UGANDA T. Kalule, M.W. Ogenga-Latigo^1 and V.A.O. Okoth^2
Namulonge Agricultural and Animal Production Research Institute P.O. Box
7084, Kampala, Uganda (Received 20 November, 1995; accepted 2 September, 1997)
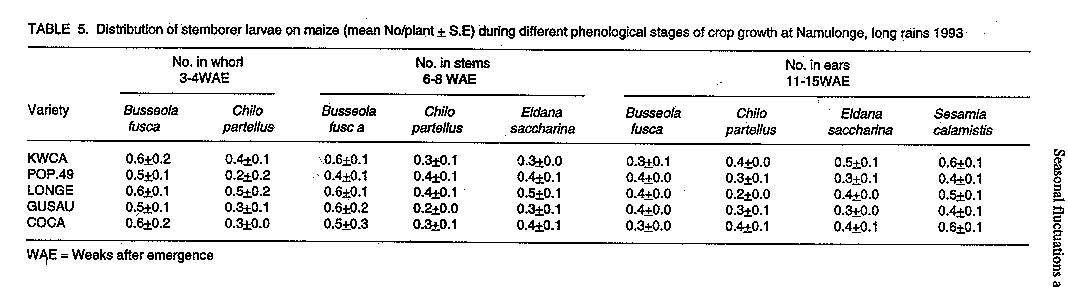
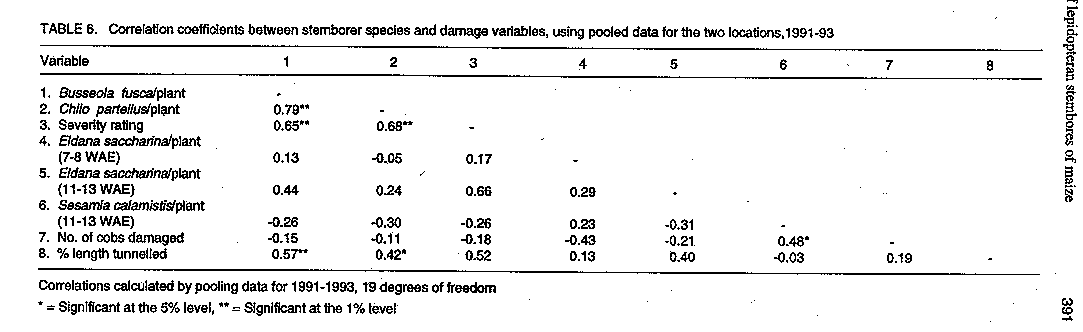
Code Number:CS97044 ABSTRACT The distribution and damage of stemborers of maize, Chilo partellus (Swinhoe), Busseola fusca (Fuller), Eldana saccharina Walker, and Sesamia calamistis Hampson, were studied in two locations in Uganda. The species composition of the stemborers was influenced by crop phenology. Distribution of stem borer larvae in the plant was similar at both locations. At 2-3 weeks after plant emergence (WAE), 100% of larvae recovered was from the funnel of the plant with the proportion decreasing as the stem elongated. Busseola fusca and Chilo partellus were more injurious as indicated by the extent of leaf and stem injury. Sesamia calamistis predominantly attacked the maize ears. This phenological relationship of variation in stemborer incidence and damage to crop phenology is crucial in devising management strategies of the pests. Key Words: Busseola fusca, Chilo partellus, Sesamia calamistis, Eldana saccharina, crop phenology, stem tunnelling, Zea mays Resume La distribution et les degets causes par les insectes terebrants (stemborers) des tiges du mais, Chillo partellus (Swinhoe), Busseola fusca (Fuller), Eldama Saccharina Walker, et Sesamia calamistis Hampson, ont ete etudies dans deux endroits en Ouganda. La composition specifique des insectes terebrants a ete influencee par la phenologie de la culture. La distribution des larves des insectes terebrants dans la plantes s'est revelee semblable dans les deux sites. Deux a trois semaines apres l'emergence de la plante (WAE), 100% des larves retrouvees provenaient de la cheminee de la plante avec une proportion decroissant avec l'elongation de la tige. Busseola fusca et Chilo partellus etaient plus agressives comme l'indiquaient l'etendue de la feuille et la blessure de la tige. Sesamia calamistis attaquait surtout les epis du mais. Cette relation phenologique de la variation dans l'incidence des insectes terebrants sur les tiges et les degets causee a la phenologie de la culture est cruciale dans les strategies combinees du controle des pestes. Mots Cles: Busseola fusca, Chilo partellus, Sesamia calamistis, Eldama saccharina, phenologie de la culture,tunnel de la tige, Zea mays. INTRODUCTION Maize (Zea mays L.) is a crop which can be grown in a wide range of environments and has multiple uses. Consequently, factors which limit its production are also diverse. Some of the most important constraints are insect pests and closely related organisms such as mites (Ortega, 1987). Lepidopteran stemborers are among the most important pests of maize in Africa. The four major species that infest maize in East Africa are the maize stalk borer, Busseola fusca (Fuller) (Noctuidae), pink stalk borer, Sesamia calamistis Hamps (Noctuidae), sugar cane borer, Eldana saccharina Walker (Pyralidae), and the spotted stalk borer, Chilo partellus (Swinhoe) (Pyralidae) (Girling, 1978). Feeding by borer larvae in whorls of the maize plant usually leads to dead hearts and early leaf senescence. After the larvae have penetrated the stems, they create tunnels by eating through the vascular bundles, leading to reduced translocation of nutrients and assimilates. Stem tunnelling also reduces plant vitality and the grain filling process, and promotes breakage of plants as they mature. The feeding activity also extends to the maize cob causing substantial damage (Bosque-Perez et al., 1987; Bosque-Perez and Mareck, 1991). A crucial factor influencing the nature of control measures applied against stemborers is the change in population of the pest over time, or its population dynamics (Seshu Reddy et al., 1990). Studies on the population dynamics and infestation patterns of stemborers, and the nature of damage, in relation to the phenological stages of the host crop are essential. They provide reliable estimates of present and future insect damage, therefore, help in making decisions as to the necessity of control (Graham and Stark, 1954). In Uganda, although damage by lepidopteran stemborers has long been recognised, few attempts have been made to ascertain the relative importance of the different borer species on maize. There is also lack of information on their population dynamics. This study was instituted to monitor natural infestations of maize by various stemborer species during the growing season, and to determine their distribution and damage in selected areas in Uganda. This information was gathered as part of effort to develop an integrated pest management programme for these insects. MATERIALS AND METHODS Field experiments were conducted at the Technology Verification Centre (TVC) at Masaka in south-western Uganda, during the long and short rains of 1991, and at Namulonge Agricultural and Animal Production Research Institute (NAARI) in southern Uganda. Trials at Namulonge were conducted during the long and short rains of 1992 and during the long rains of 1993. These areas occupy the southern and western tall grassland zone and extend into the pastoral dry to semiarid rangeland zone of western Masaka. Both areas are found in the mid-altitude zones of Uganda at 1000-1500 metres above sea level. At Masaka planting was in April in the long rains and late September in the short rains, while at Namulonge the planting was in late March in the long rains, and September in the short rains. Five maize varieties of varying maturity periods, namely Longe 1, GUSAU, KWCA-Certified, COCA and Population 49, were used. Their duration to 50% silking were 68, 67, 78, 68 and 56 days, respectively. Sampling blocks were established by sowing the five maize varieties in adjacent plots replicated four times. The plots measured 5.5 x 3.75m, each consisting of four 5m long rows, with maize planted at a spacing of 75cm between rows and 50cm within the rows. Three seeds were planted per hole and the seedlings thinned to two plants per hill two weeks after planting. Triple superphosphate fertilizer was applied at planting at a rate of 60 kg P2 05 ha^-1. Nitrogen was applied at a rate of 90 kg ha^-1 as urea 3-4 weeks after planting. Weeding was carried out at 2, 5 and 7 weeks after crop emergence (WAE). The incidence and species composition of the lepidopterous stemborers of maize were established by randomly selecting 10 maize plants from each plot at two week intervals from the date of germination up to harvest. First instar larvae were counted by careful examination of the whorl. Older larvae and pupae were counted by randomly uprooting ten plants per plot and splitting each open along its length to expose the insects. Maize cobs were also examined for the presence of larvae. Borer species recovered were identified on the basis of body pigmentation and the abdominal crotchets (Girling, 1978). Pupae recovered were allowed to emerge for identification of species at adult stage. Stemborer damage severity on maize whorl leaves was assessed by taking damage scores based on the amount of feeding from four uppermost leaves. A scale of 1-5 was used, whereby 1= 1-20%; 2=21-40%; 3= 41-60%; 4= 61-80%; and 5 = over 81% damage or complete death of the leaf (Anon., 1990). The extent of stemborer damage on maize stems was assessed by measuring the length of tunnels created by borer feeding on ten randomly selected plants. Tunnel length per plant was expressed as percentage using height of the stem to the topmost ear bored (Bosque- Perez and Mareck, 1991). Borer data was subjected to logarithmic transformations log(x+1) before statistical analysis and percentages of the other variables subjected to arcsine transformations. Step-wise regressions were used to determine relationships between borer species and maize damage. RESULTS All four major stemborer species, Chilo partellus, Busseola fusca, Eldana saccharina, and Sesamia calamistis, were found feeding on all the maize varieties, both at Masaka and Namulonge. Several species were occasionally found on the same plant. Marked differences were observed in the seasonal incidence of stem borers on maize among locations. In Masaka, for instance, B. fusca was the dominant species from 2-8 weeks after crop emergence in both seasons (Table 1). Its population slightly declined after the 8th week. Chilo partellus was the second most abundant species during this period (Table 1 ). Infestation by E. saccharina at Masaka commenced around 6 WAE, with peak infestations at 7-8 WAE. By harvest, E. saccharina had become the dominant borer species. Sesamia calamistis attack occurred only later, at 10-12 WAE (Table 1). At Namulonge, B. fusca exhibited the same level of incidence with C. partellus during the first 4 weeks of crop development. During the 6th and 7th week however, its population declined relative to C. partellus (Table 2). Eldana saccharina infestation at Namulonge occurred later (7- 8 WAE) as compared to 5-6 WAE at Masaka. Sesamia calamistis attack occurred late at both Namulonge and Masaka (11-12 WAE) (Table 2). The distribution of stem borer larvae within the plants at Masaka was similar in both seasons, with the majority of larvae being found in the stems (Table 3). Busseola fusca and C. partellus larvae were found as early as 2-3 WAE, and were the only larvae attacking whorl leaves of maize. At this stage, almost 100% of the larvae recovered were found in the funnel of the maize plants. The proportion of the larvae in the funnel decreased as the stem elongated. By 6-8 WAE, most of the larvae were recovered from stems, with the predominant species being B. fusca and C. partellus followed by E. saccharina. Sesamia calamistis occurred predominantly in the ears (Table 3). A similar trend was observed at Namulonge (Table 4). At this stage it was common to find B. fusca and C. partellus in the stem of the same plant. In both locations, no clear relationship was obtained between cultivar and stemborer preference. However, E. saccharina and S. calamistis, both infesting the crop at late growth stages, were more predominant at 11-15 WAE on the late maturing variety, KWCA, than on the early maturing Population 49 (Tables 3 and 4). In the long rains of 1993, proportionately higher numbers of B. fusca were recovered from the leaf whorl and in the stem than those of C. partellus (Table 5). A correlation matrix relating the various pest and damage variables calculated from the pooled data of 1991-93 is presented in Table 6. Both B. fusca and C. partellus showed a positive relationship with severity of leaf damage. Busseola fusca, C. partellus and E. saccharina numbers were positively correlated to stem tunnelling although the latter was not significant. Sesamia calamistis, unlike the other borers, was positively correlated to the number of cobs damaged (Table 6). The relative importance of B. fusca (x1) and C. partellus (x2) on severity of leaf damage (Y), was described by the equation; Y= 1.0 + 0.81 x 1 + 0.67 x2, r^2= 0.56. Both species significantly influenced the severity of leaf damage (t-values for x1 and x2 were 2.1* and 2.3*, respectively) with B. fusca accounting for more damage than C. partellus (0.81 and 0.67, respectively). A multiple regression relating stem tunnelling (Y), with borer species, B. fusca (x1), C. partellus (x2), and E. saccharina (x3), showed that only B. fusca and C. partellus were significantly correlated with stem tunnelling Y= 1.6 + 2.9 x 1 -1.3 x 2 + 0.2 x 3, r^2 = 0.60, suggesting that tunnel damage was mainly caused by these pests. (t-values x1, x2, x3 were 4.1**, -1.8* and 0.1, respectively). When S. calamistis (x1), and E. saccharina (x2) populations were regressed on cob damage, only S. calamistis was of importance (Y =1.5 + 1.3x1 -1.0x2 ,r2 = 0.48; t-values for x1, x2 were 2.4* and -0.8, respectively). DISCUSSION The present study confirmed that C. partellus, B. fusca, E. saccharina and S. calamistis cause damage to maize in Uganda as reported earlier by Girling (1978). Busseola fusca and C. partellus were associated with the crop throughout the season while E. saccharina attacked just before anthesis stage and S. calamistis was a post anthesis pest. Ingram (1958) reported that C. partellus could not live above 1200m in areas of southern and western Uganda. On the other hand, Nye (1960) found B. fusca to be the dominant borer in the mainly cool and high altitude zones above 1050m. Our observations, however, indicate that this distribution appears to have changed. Chilo partellus is now a common borer above 1200m in the dry and warm areas of Masaka. In addition, B. fusca is also well established in the moist and humid mid altitude (1000-1500 m.a.s.l) areas of southern Uganda. The surveys by Ingram (1958) and Nye (1960) had also established that E. saccharina was a pest of minor importance on maize and was restricted to the sugar belts. In this study, however, this pest was observed in substantial proportions feeding on maize stems and cobs in the banana-maize cropping agroecozones of Namulonge and Masaka. Eldana saccharina, therefore, appears to have widened its distribution since then. These changes could be attributed to changes in cropping patterns from a predominantly banana based to a banana-maize cropping system. The study further showed that the level of borer attack varied with the phenological stages of the crop and their preferences for the crop. While B. fusca and C. partellus attacked the crop from seedling stage until harvest, E. saccharina attacked maize during the late vegetative stage, while S. calamistis attacked mainly the cobs. This is in agreement with observations made by Bowden (1959) and Girling (1978). The decrease in numbers of B. fusca and C. partellus during the late stages of the crop could have been due to a decrease in larval density per plant and increase in the number of diapausing larvae, while higher numbers of E. saccharina and S. calamistis could be attributed to the abundance of young instar larvae at post anthesis stage. Stem tunnelling is a good indicator of the degree of plant damage and, thus, yield loss. The more extensive the tunnelling, the higher the yield loss (Bosque- Perez and Mareck, 1991; Kalule et al., 1994). Van den Berg and Van Rensburg (1991) also found considerable yield loss in sorghum due to stem tunneling by C. partellus. In our study, B. fusca and C. partellus caused extensive tunnelling to the plant in the early stages of development. This, therefore, underlines the importance of these borers to the maize crop. The studies reported here have confirmed the occurrence of the stemborer complex on maize in varied densities in southern Uganda. The study also showed that the stemborer species occupy different niches on the plant and hence have different damage patterns. There is, therefore, need to develop appropriate management strategies for stemborers of maize in Uganda based on their importance and behaviour on the crop. ACKNOWLEDGEMENTS Financial assistance was provided by the United States Agency for International Development (USAID) under the Manpower for Agricultural Development Program (MFAD). REFERENCES Anonymous. 1990. Annual Report of Namulonge Research Station. Ministry of Agriculture, Animal Industry and Fisheries, Uganda, pp. 143-144. Bosque-Perez, N.A. and Mareck, J.H. 1991. Effect of the stem borer Eldana saccharina (Lepidoptera) on the yield of maize. Bulletin of Entomological Research 81:243-247. Bosque-Perez, N.A., Mareck, J.H., Dabrowski, Z.T., Everett, L., Kim, S.K. and Efron, Y. 1987. Screening and breeding maize for resistance to Sesamia calamistis and Eldana saccharina. In: Towards Insect Resistance in Maize for the Third World: Proceedings of the International Symposium on the Metho-dologies for Developing Host Plant Resistance to Maize Insects. pp. 163-169. Mexico, D.F. CIMMYT. Bowden, J. 1959. The stem borer problem in tropical cereal crops. Report of the 6th Commonwealth Conference. pp. 107-134. Girling, D.J. 1978. The distribution and biology of Eldana saccharina Walker (Lepidoptera: Pyralidae) and its relationship to other stem borers in Uganda. Bulletin of Entomological Research 68:471-488. Graham, K. and Stark, R. W.1954. Insect population sampling. Report of Fifty Third Annual Meeting of the Entomological Society, British Columbia, pp.1-6. Ingram, W.R. 1958. The lepidopteran stalk borers associated with graminae in Uganda. Bulletin of Entomological Research 49:367-383. Kalule, T., Ogenga-Latigo, M.W. and Okoth, V.A.O. 1994. Relationship between successive dates of sowing maize and stemborer damage in central Uganda. Proceedings of African Crop Science Conference 1:338-340. Nye, I.W.B. 1960. The Insect Pests of Gramin-aceous Crops in E. Africa. Colonial Research Studies. No. 31. London HMS. Ortega, A. 1987. Insect Pests of Maize. A Guide to Field Identification. CIMMYT, Mexico, D.F. Seshu-Reddy, K.V., Lubega, M.C. and Sum, K.O.S. 1990. Population patterns of Chilo spp. in sorghum, maize and millets. Insect Science and its Application 11:549-559. Van den Berg, J. and Van Rensburg J.B.J. 1991. Comparative injuriousness of Busseola fusca (Lepidoptera: Noctuidae) and Chilo partellus (Lepidoptera: Pyralidae) on grain sorghum . Bulletin of Entomological Research 82:137-142.
Copyright 1997, African Crop Science Society The following images related to this document are available:Line drawing images[cs97044b.gif] [cs97044a.gif] [cs97044e.gif] [cs97044c.gif] [cs97044f.gif] [cs97044d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}