 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
ANTIBIOSIS AND ANTIXENOSIS OF TWO COWPEA VARIETIES TO THE LEGUME FLOWER THRIPS S. EKESI, N.K. MANIANIA and I. ONU^1
International Centre of Insect Physiology and Ecology, P.O. Box 30772,
Nairobi, Kenya (Received 9 April, 1997; accepted 16 October, 1997)
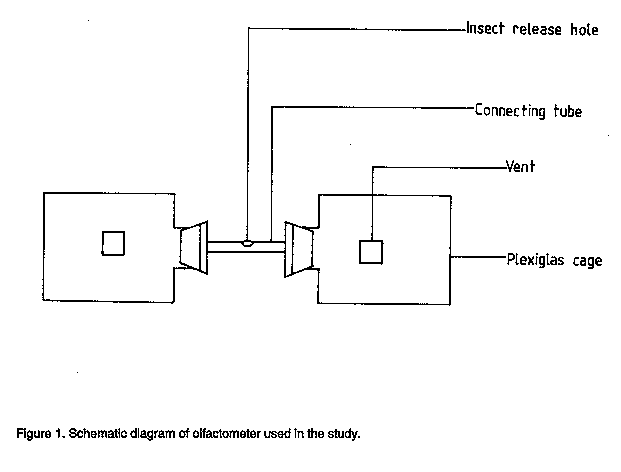
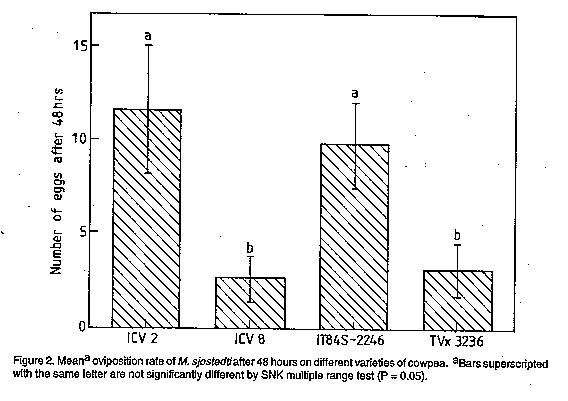
Code Number:CS98006 ABSTRACT Laboratory investigations were conducted to study the mechanisms of resistance of two cowpea (Vigna unguiculata L. Walp) varieties, ICV 8 and IT84S-2246 to the legume flower thrips, Megalurothrips sjostedti (Trybom) [Thysanoptera: Thripidae]. High mortality of larval stages and slower developmental rates on ICV 8, indicated presence of antibiotic mechanisms of resistance. Adult longevity and oviposition rates of thrips fed on pollen of ICV 8 were greatly reduced. Mean larval and pupal weights were also low on this variety but adult weight was not affected. Mortality rates on IT84S-2246 was low and developmental rates was normal, indicating an absence of antibiotic mechanisms. However, a choice experiment using cowpea flowers, and olfactometer assessment, showed that IT84S-2246 was the least preferred variety, indicating evidence of antixension as the mechanism of resistance in this variety. Key Words: Megalurothrips sjostedti, resistance, olfactometer, Vigna unguiculata RESUME Des etudes ont ete conduites au laboratoire pour etudier le mecanisme de resistance de deux varietes de niebe (Vigna unguiculata L. Walp), ICV 8 at IT84S-2246, a l'egard du thrips des fleurs de niebe, Megalurothrips sjostedti (Trybom) (Thysanoptera: Thripidae). Une mortalite elevee des stades larvaires ainsi qu'un long developpement larvaire ont ete observes avec la variete ICV 8, montrant ainisi la presence des mecanismes antibiotiques de resistance. La longevite des adultes et le taux d'oviposition des insectes nourris au pollen de la variete ICV 8 etaient considerablement reduits. Les poids moyen des larves et des pupes etait egalement peu eleve avec cette variete; toutefois, le poids des adultes n'etait pas affecte. Avec la variete IT84S-2246, la mortalite etait peu elevee, tandis que le taux de developpement larvaire etait normal, indiquant ainsi l'absence des mecanismes antibiotiques. Cependant, un test a double choix avec les fluers de niebe et un essai dan un olfactometre, ont revele que la variete IT84S-2246 etait la moins preferee, indiquant ainsi que l'antixenose est le mecanisme de resistance pour cette variete. Mot Cles: Megalurothrips sjostedti, resistance, olfactometre, Vigna unguiculata INTRODUCTION The legume flower thrips, Megalurothrips sjostedti (Trybom) is a pest of economic importance on cowpea, Vigna unguiculata (L.) Walp in tropical Africa. Damage by the pest begins at the terminal leaf bud stage of the cowpea plant and spreads to flower buds and flowers leading to necrosis and/or abscission (Ezueh, 1981). Yield losses attributed to this pest range from 20-100% in different parts of Africa (Singh and Allen, 1980). Current control tactics include mainly the use of chemical insecticides (Singh and Taylor, 1978; Afun et al.,1991). This method is, however, associated with various ecological problems. Consequently, attention has been directed towards the use of resistant varieties which are likely to be more environmentally benign and compatible with other control measures. A thorough understanding of insect-plant interaction on different varieties is an important prerequisite to efficient utilisation of resistance in a pest management programme (Kennedy, 1978). Also the knowlegde of resistance mechanisms on different varieties is essential for providing useful guidelines for selection and incorporation of appropriate resistance genes into different varieties. The influence of host plant resistance on various thrips species have been studied (Stoner and shelton, 1988; Mollema et al., 1993; De Jager et al., 1995). The mechanism of cowpea resistance to M. sjostedti has, however, received little attention. The only extensive study of resistance mechanism of cowpea to M. sjostedti was done by Salifu et al. (1988a,b) on TVx 3236 and by Rosingh (1980) on TVu 1509 which is a parent variety of TVx 3236. Further detailed studies on different varieties of cowpea are required. Host plant resistance is often the result of a combination of resistance categories. Assessment of both antibiotic and antixenotic mechanisms of resistance are, therefore, important in identifying the effect of each category of resistance to the overall expression of resistance (Van Emden, 1987). In the present study, resistance mechanisms of two cowpea varieties, ICV 8 and IT84S-2246 widely grown by farmers, and which have been reported to have less thrips in field samples (Pathak and Olela, 1986; IITA, 1987), indicating possible display of antibiosis or antixenosis was examined. Antibiosis was tested by studying the effect of the different cowpea varieties on development, fecundity and mortality of M. sjostedti. Antixenosis was observed in a choice-experiment using cowpea flowers and in an olfactometer. MATERIALS AND METHODS Plant and insect culture. Four cowpea varieties: ICV 2, ICV 8, IT84S-2246 and TVx 3236 were used for the various experiments. ICV 2 and ICV 8 comprise varieties developed and registered at the International Centre of Insect Physiology and Ecology (ICIPE) (Pathak and Olela, 1986) while IT84S-2246 and TVx 3236 are advanced lines developed at the International Institute of Tropical Agriculture (IITA). ICV 2 has a spreading, indeterminate growth habit and matures in 60 days. It is very susceptible to M. sjostedti but has good tolerance to pod bugs and pod borers and was used as a susceptible check. ICV 8 is semi-erect, determinate and matures in 60 days. It is moderately resistant to M. sjostedti but the mechanism of resistance is not known. IT84S-2246 is erect, determinate and matures in 65 days. It is resistant to aphids and storage bruchids and moderately resistant to flower thrips, but the mechanisms of resistance to thrips has not been investigated (IITA, 1987; Afun et al.,1991). TVx 3236 is semi-erect, determinate, and moderately resistant to flower thrips and matures in about 75 days. It has antibiotic and antixenotic mechanisms of resistance to M. sjostedti (Salifu et al.,1988a,b) and was used as a resistance check. A culture of M. sjostedti was maintained following modification of Mollema et al.(1995) in a controlled room environment at 26 +/-20 C and 60 +/-5% RH under a photoperiod of L:D 12:12. The initial stock culture originated from natural population of M. sjostedti infesting a local variety of cowpea, Ex-Luanda, at the ICIPE Headquarters, Nairobi, Kenya. Adult female thrips were aspirated from cowpea flowers and transferred to 1 litre glass jar containing Ex-Luanda cowpea pods as oviposition substrate. Pollen was dusted from the anthers of cowpea flowers on the pods. Megalurothrips sjostedti feed on pollen and on the skin of the pods, which also serve as oviposition substrate. Glass jars were covered with microscreens with a mesh size of 192 um. Female thrips were allowed to oviposit for one day, after which time the pods were brushed off and transferred to another jar. First instar larvae emerged on the surfaces of pods within 2-3 days. Larvae were reared on pods until they were about to pupate. They were then transferred to screened transparent plastic vials (50 x 100mm) which were lined with moistened cheesecloth and sandwiched between two layers of filter paper. Pupation occurred on the underside of the first and second layer of filter paper. Adults emerged within 4-5 days and, were used for the various experiments. Antibiosis. Six adult female M. sjostedti were collected at random from the culture and placed in polythene specimen tubes (95 x 30mm) containing upper 5cm of cowpea peduncles of individual variety under test, as oviposition substrate. Although, M. sjsotedti oviposit on leaf petioles, calyx tubes and pods, peduncles have been reported as the preferred oviposition substrate (Tamo et al., 1993). To obtain synchronised first instar larvae, females were allowed to oviposit for one day, after which they were carefully removed. Three days later, 30 newly emerged larvae were placed individually on floral tissues of each variety which was changed daily. Record of developmental stages were kept for each larva. Transmission from one stage to another was distinguished by microscopic examination for exuviae left on floral tissues. The weight of larva II, pupa and adult were also taken at each stage. Larval mortality was observed under a similar set up using 20 larvae per variety. These experiments were replicated three times. ANOVA procedure and Student-Newman-Keuls (SNK) multiple range test of SAS (SAS Institute, 1989) were used to test for differences in developmental time and mortality between different varieties. Adults which emerged from the above experiments were used to test for oviposition and adult longevity. Three-day old adult female thrips each emerging from the four varieties under test and which had been previously paired with a male thrip was placed in a cage (60 x 30mm). One end of the cage was covered with a stretched Parafilm membrane. A drop of distilled water was deposited on the stretched parafilm membrane and covered with another layer of stretched parafilm membrane. Eggs were deposited in the distilled water between the double layer of stretched parafilm membrane. Thrips were fed on pollen from each of the four varieties under test. Egg counts were taken after 48hr. The experiment was replicated five times. Adult longevity was determined by retaining newly emerged adult thrips on flowers of each of the four varieties in a cage (40 x 20 mm). Flowers were changed daily and longevity was recorded as the time from emergence until death on the different varieties. The exepriment was replicated four times. ANOVA procedure and SNK multiple range test of SAS were used to test for differences in oviposition and adult longevity between the varieties. Antixenosis. This was assessed under a choice situation using cowpea flowers. Seeds of the four cowpea varieties were sown in 0.85 litre plastic pots filled with pasteurised soil. Planting was scheduled in such a way that flowering of all varieties occurred simultaneously. Four flowers, one from each of the four varieties were picked from the plant and arranged at equal distance around the edge of a transparent circular plastic box (30 x 10cm) covered with microscreen. Fourty adult thrips were aspirated into a vial from the culture. They were then immobilised by chilling them for 25 seconds and released at the centre of the box. Twenty four hours later, test boxes were opened and the flowers were placed in a container of 40% alcohol. The number of thrips on each flower was counted and recorded. The experiment was replicated five times and the data were analysed using the ANOVA procedure and SNK multiple range test of SAS. Olfactory assay. The straight-tube olfactometer used (Fig.1) is similar to that described by Kyamanywa et al. (1993). It consisted of two plexiglas cage (12 x 12 x 12cm) used for the odour source and a connecting tube with a hole for introduction of insects. The cages are connected in such a way that insects introduced into the tube could not see the odour source, therefore, movement towards or away from the cages were associated only with the odour which spread by passive diffusion. Treatment combinations included: Blank vs blank, susceptible variety vs blank, resistant varieties vs blank, susceptible vs resistant varieties and resistant vs resistant varieties. Twenty adult insects were obtained from the culture, immobilised as described earlier and introduced into the olfactometer using a camel hair brush. The response of insects to odour sources was recorded after one interval of 15 minutes by counting the number of thrips that moved to either sides of the cage and the number not responding to odour source. Twenty cowpea flowers from representative varieties, or blanks in two choice combinations were tested. To avoid any effect of position of olfactometer on thrips response, the system was rotated after every five minutes. Tubes and cages were washed with detergent and rinsed thoroughly in distilled water at the end of each test. All tests were conducted at 26 +/-20C and 60 - 70% Relative humidity in an area provided with a photographic red light at the level of experimental insects. Five readings were made for each test and data obtained were analysed using Mann-Whitney U-test. RESULTS AND DISCUSSION Antibiosis. Results of developmental time is presented in Table 1. Significant difference (P < 0.05) was observed in total developmental period between different varieties. This was longer on ICV 8 compared to IT84S-2246 and the susceptible check, ICV 2. There was no significant difference between ICV 8 and the resistance check, TVx 3236. Developmental period on IT84S-2246 did not differ significantly from the susceptible check, ICV 2. The duration of egg, larva I, prepupa and pupa on the different varieties were not significant. However, significant difference (P < 0.05) occurred in larva II among the various varieties. Differences in total developmental time was, therefore, due to prolonged developmental period of larva II on the resistant varieties. Salifu et al. (1988a,b) and Mollema and Soria (1995) have reported similar observation on M. sjostedti and Frankliniella occidentalis (Pergande) when reared on resistant lines of cowpea and cucumber, respectively. Prolonged developmental period on resistant varieties could be due to reduced food intake due to toxins, malnutrition or deterrents (Bakker, 1961). These factors adversely affects thrips larval stages as reported by Trichilo and Leigh (1988). Extension of immature period could reduce the number of generations, increasing the effects of predation parasitism and disease (Painter, 1951). Table 2 shows the weight of the various developmental stages on different varieties. Larvae and pupae reared on IT84S-2246 were significantly heavier than those reared on ICV 8 and TVx 3236. There was no significant difference between the weights of larvae and pupae reared on IT84S-2246 and the susceptible check, ICV 2. However, contrary to expectations, adult emerging from pupae on ICV 8 which produced low pupal weights, did not significantly produced lighter adults than those emerging from pupae on ICV 2 and IT84S-2246. Although Salifu et al. (1988b) did not find any significant difference in adult weights reared on resistant and susceptible varieties, our results indicate a strong effect of resistant varieties on weights of different developmental stages. The effect of resistant varieties on larval, pupal and adult weights could be explioted by incorporation into microbial and chemical control programmes. This combination may become more efficacious as the susceptibility of the insect to either insecticide is increased owing to reduced body weight and/or physiological stress imposed on the insect as a result of feeding on suboptimal hosts (Hare, 1992). Larval mortality was considered one indicator for the presence or absence of antibiosis on various varieties. Mortalities in larva I and II on ICV 8 averaged 21% and 56%, respectively (Table 3). This did not differ significantly from mortalities on the resistance check, which averaged, 23% and 51% for larva I and II, respectively. On IT84S-2246 mortality was low with larva I and II incurring 13% and 17% mortality, respectively. This did not differ significantly from the susceptible check, which averaged 10% and 17%, respectively. Generally, mortality in larva II was higher compared to larva I. This is similar to observations of Soria and Mollema (1995) and these authors attributed low mortality in the first instar larva of F. occidentalis on cucumber to nutritive reserves sustained from egg. Adult logevity was significantly shorter (P < 0.05) on resistant lines compared to the susceptible line. This concur with observation of Salifu et al. (1988b). These authors reported shorter adult longevity on resistant TVx 3236 compared to the susceptible VITA 7. Megalurothrips sjostedti reared on ICV 8 that were able to develop to adulthood produced significantly (P < 0.01) fewer eggs when compared to thrips reared on IT84S-2246 and the susceptible check, ICV 2 (Fig.2). There was no significant difference between ICV 8 and the resistance check, TVx 3236. Pollen was the only source of food provided during the period of oviposition. Egg production is known to be affected by the amount of protein in an insect diet (Engleman, 1984) and pollen, which is known to be high in protein (Mound et al., 1980) is an important requirement of egg production in thrips (Trichilo and Leigh, 1988; Teulon and Penman, 1991). The results obtained, therefore, indicate true difference in oviposition as affected by diet from the different varieties. Nutritional differences between species of pollen have been reported for many insects (Pajni and Sood, 1975; Wahl and Ulm, 1983; Kirk 1985a). Soria and Mollema (1995) suggested that exposure of female thrips to toxins, deterrents or antifeedants on resistant cultivars may reduce food intake and subsequently affect egg production. Moreover, poor oviposition as a result of inadequate nutrition are also well established for thrips (Laughlin, 1971; Murai and Ishii, 1982; Kirk, 1985a). Antixenosis. Significantly (P < 0.05) fewer thrips were recovered from flowers of IT84S-2246 compared to other varieties (Fig.3). The susceptible check attracted more thrips compared to other varieties. There was, however, no significant difference between the number of thrips recovered from ICV 8 and TVx 3236. The rice thrips, Stenchaetothrips biformis Bagnall, rarely colonise resistant varieties of rice in a choice-experiment (Parthiban and Veeravel, 1994). Stoner and Shelton (1988) also reported reduced number of Thrips tabaci Lindeman on resistant varieties of cabbage compared to susceptible lines. Results from the olfactory assay are presented in Table 4. Assessment of blank vs blank treatment revealed no significant difference. ICV 2, ICV 8 and TVx 3236 attracted more thrips (61%, 56% and 51%, respectively) than the blank controls (13%, 21% and 17%, respectively). Significantly fewer thrips (19%) were attracted to IT84S-2246 compared to the blank control (45%). The highest percentage of non-responding thrips when all varieties were tested against blank occurred in IT84S-2246 vs blank. When resistant varieties were tested against the susceptible variety, significantly more thrips were attracted to ICV 2 compared to other varieties except when tested against ICV 8. There was no significant difference when ICV 8 was tested against TVx 3236. ICV 8, however, attracted more thrips (56%) than IT84S-2246 (17%) when both varieties were tested. Results of TVx 3236 vs IT84S-2246 revealed no significant difference. This test also produced the highest percentage of non-responding thrips among the resistant vs resistant entries. The results of the olfactory assay showed that M. sjostedti responded to odour of host plant. Kyamanywa et al. (1993) found that high number of M. sjostedti oriented towards sole cowpea odour source than cowpea/maize odour source. Host selection by insects is mediated by visual and olfactory cues (Visser, 1986). Although the host-finding cues of M. sjostedti is not known, Kirk (1985b) suggested odour-induced visual response for thrips. Adult thrips are weak flyers (Lewis, 1973; Ananthakrishnan, 1984) and flight is based on drag rather than lift (Nachtigall, 1989). Teulon (1993), therefore, suggested that host-finding responses could also be based on odour-induced olfactory stimuli or chemotaxis. The strong response observed in the olfactometer may probably be due to this conditions. Steep gradients of odour concentration in an olfactometer have been reported to permit strong response of various insects to odour sources (Kennedy, 1977). Expression of antixenosis through host plant odour have also been reported for many insects. Gaum et al. (1994) reported that F. occidentalis showed different response to odour source of rose cultivars in an olfactometer. Similarly, Storer and Van Emden (1995) showed that Aphis gossypii Glover showed different response to host plant odour from different cultivars of chrysanthemum when tested in an olfactometer. Steam distillate extracts of volatiles from resistant and susceptible cultivars of rice have been reported to influence the behaviour of whitebacked Sogatella furcifera (Horvath) and brown plant hopper, Nilaparvata lugens (Stil) (Saxena and Okech, 1985; Khan and Saxena, 1986). Furthemore, extracts from different cultivars of cowpea has been shown to either attract or repel cowpea curculio (Chalcodennus aeneus) in an olfactometer (Rymal et al., 1981). Although it was realised that all resistant varieties tested in this studies were indeed resistant to M. sjostedti, this investigation succeeded in identifying the categories of resistance in ICV 8 and IT84S-2246 which were previously not known. Resistance in ICV 8 is due mainly to antibiosis as observed by poor development of thrips on this variety. Oviposition rates on this variety were also low, which suggest that poor nutrition from ICV 8 had significant effect on oviposition. The resistance of IT84S-2246 appears to be antixenotic. The lack of statistically significant difference in developmental period between this variety and the susceptible line shows that antixension rather than antibiosis is involved in the resistance. The high "repellency" observed in the laboratory test with cowpea flowers of IT84S-2246 and in the olfactometer provide additional evidence of antixension in this variety. TVx 3236 combines both antibiosis and antixenosis which is parallel to findings of Salifu et al. (1988 a,b). Further studies should concentrate on identification of factors which affect development and behaviour of the pest on these varieties. This will provide plant breeders with valuable information for the development of varieties of cowpea resistant to M. sjostedti. ACKNOWLEDGEMNTS The authors would like to thank Dr. M.E. Muhinda (ICIPE) for reviewing the manuscript. We also thank Dr. A. Odulaja (ICIPE) for statistical advice and Mr. E. Wesonga for technical assistance. The research was funded by ICIPE. REFERENCES Afun, J.V.K.,Jackai,L.E.N. and Hodgson, C.J. 1991. Calendar and monitored insecticide application for the control of cowpea pests. Crop Protection 10:363-367. Ananthakrishnan, T.N. 1984. Bioecology of thrips. Indira. Oak Partk, IM. Bakker, K. 1961. An analysis of factors which determine success in competition for food among larvae of Drosophila melanogaster. Archives Neerlandaises de Zoologie 14:200-281. De Jager, C.M., Butot, R.P.T., Klinkhamer, P.G.L., de Jong, T.J., Wolff, K. and Meijden, E. van der. 1995. Genetic variation in chrysanthemum for resistance to Frankliniella occidentalis. Entomologia Experimentalis et Applicata 77: 277-287. Englemann, F. 1984. Reproduction in insects. In: Ecological Entomology. Hufferker, C.B. and Rabb , R.B.(Eds.), pp.113-114. John Wiley and Sons, New York. Ezueh, M.I. 1981. Nature of preflowering damage by thrips to cowpea. Entomologia Expe-rimentalis et Applicata 29:305-312. Gaum, W.G., Giliomee, J.H. and Pringle, K.L. 1994. Resistance of some rose cultivars to the Western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Bulletin of Entomological Research 84:487-492. Hare, J.D. 1992. Effects of plant variation on herbivore-natural enemy interactions. In: Plant resistance to herbivores and pathogens: ecology, evolution, and genetics. Fritz, R.S. and Sims, E.L. (Eds.), pp. 278-298. University of Chicago Press, Chicago. IITA, (International Institute of Tropical Agriculture) 1987. Annual Report for 1986. IITA, Ibadan, Nigeria. Kennedy, J.S. 1977. Olfactory response to distant plants and odour sources. In: Chemical Control of Insect Behaviour. Shorey, H.H. and McKelvy, J.T. (Eds.), pp. 67-91. John Wiley & Sons, London. Kennedy, G.G. 1978. Recent advances in insect resistance of vegetables and fruit crops in North America. Bulletin of the Entomological Society of America 24:375-384. Kirk, W.D.J. 1985a. Pollen-feeding and the host specificity and fecundity of flower thrips (Thysanoptera). Ecological Entomology 10: 281-289. Kirk, W.J.D. 1985b. Effect of some floral scents on host finding by thrips (Insecta: Thysa-noptera). Journal of Chemical Ecology 11: 35-43. Khan, Z.R. and Saxena, R.C. 1986. Effect of steam ditillate extracts of resistant and susceptible rice cultivars on behavoiur of Sogalella furcifera (Homoptera: Delp-hacidae). Journal of Economic Entomology 79:928-935. Kyamanywa, S., Baliddawa, C.W. and Ampofo, J.K.O. 1993. Effect of maize plants on colo-nisation of cowpea plants by bean flower thrips, Megalurothrips sjostedti. Entomologia Experimentalis et Applicata 69:61-68. Laughlin, R. 1971. A culture method for Hercinothrips femoralis (Reuter) (Thys-anoptera). Journal of the Australian Entomological Society 10:301-303. Lewis, T. 1973. Thrips-their biology, ecology and economic importance. Academic Press, London, 349pp. Mollema, C., Steenhuis, G. and Inggamer, H. 1995. Genotypic effect of Cucumber res-ponse to infestation by Western Flower thrips. In: Thrips Biology and Management. Parker, B.L., Skinner, M. and Lewis, T. (Eds.), pp. 397-401. Plenum Press, New York. Mound, L.A., Heming, B.S. and Palmer, J.M. 1980. Phylogenetic relationships between the families of recent Thysanoptera (Insecta). Zoological Journal of the Linnean Society 69: 111-141. Murai, T. and Ishii, T. 1982. Simple rearing method for flower thrips (Thysanoptera: Thripidae) on pollen. Japanese Journal of Applied Entomology and Zoology 26:149-154. Nachtigall, W. 1989. Mechanics and aerodynamics of flight. In: Insect Flight. Golgsworthy, G.J. and Wheeler, C.H. (Eds.), pp.1-29. CRC, Boca Raton, FL. Painter, R.H. 1951. Insect Resistance in Crop Plants. MacMillan & Co., New York, 520pp. Pajni, H.R. and Sood, S. 1975. Effect of pea pollen feeding on maturation and copulation in the beetle, Bruchus pisorum L. Indian Journal of Experimental Biology 13:202-203. Parthiban, M.P. and Veeravel, R. 1994. Screening rice accensions for resistance to thrips. Inte-rnational Rice Research Notes 19:14-15. Pathak, R.S. and Olela, J.C. 1986. Registration of 14 cowpea cultivars. Crop Science 26:647-648. Roesingh, C. 1980. Untersuchungen uber die Resistenz edr Kuherbse, Vigna unguiculata (L.) Walp gegen Megalurothrips sjostedti (Trybom) (Thysanoptera: Thripidae). Ph.D thesis, Universitat Hohenheim. 64pp. Rymal, K.S., Chambliss, O.L. and McGuire, J.A. 1981. Role of volatile principles in non preference resistance to cowpea curculio in southern pea, Vigna unguiculata (L.) Walp. HortScience 16:670-672. Salifu, A.B., Hodgson, C.J. and Singh, S.R. 1988a. Mechanism of resistance in cowpea (Vigna unguiculata (L.) Walp) genotype, TVx 3236, to the Bean Flower Thrips, Megalurothrips sjostedti (Trybom) (Thysanoptera: Thripidae) 1. Ovipositional nonpreference. Tropical Pest Manangement 34:80-184. Salifu, A.B., Singh, S.R. and Hodgson, C.J. 1988b. Mechanism of resistance in cowpea (Vigna unguiculata (L.) Walp) genotype, TVx 3236, to the bean flower thrips, Megalurothrips sjostedti (Trybom) (Thysanoptera: Thripidae). 2. Nonpreference and antibiosis. Tropical Pest Management 34:185-188. SAS Institute. 1989. SAS user's guide: statistics. Version 6.06 Ed. SAS Institute, Cary, N.C. Saxena, R.C. and Okech, S.H. 1985. Role of plant volatiles in resistance of selected rice varieties to brown plant hopper, Nilaparvata lugens (StŒl) (Homoptera: Delphacidae). Journal of Chemical Ecology 11:1601-1616. Singh, S.R. and Taylor, T.A. 1978. Pests of grain legumes and their control in Nigeria. In: Pests of Grain Legumes: Ecology and Control. Singh, S.R., Emden, H.F. van and Taylor, T.A. (Eds.), pp. 99-111. Academic Press, London. Singh, S.R. and Allen, D.J. 1980. Pests, diseases, resistance and protection in cowpeas. In: Advances in Legume Science. Summerfield, R.J. and Bunting, A.H. (Eds.), pp. 419-443. Kew, Royal Botanic Gardens and Fisheries, MAFF, London. Soria, C. and Mollema, C. 1995. Life-history parameters of Western flower thrips on susceptible and resistant cucumber genotypes. Entomologia Experimentalis et Applicata 74: 177-184. Stoner, K.A. and Shelton, A.M. 1988. Role of nonpreference in the resistance of cabbage varieties to the onion thrips (Thysanoptera: Thripidae). Journal of Economic Entomology 81:1062-1067. Storer, J.R. and Van Emden, H.F. 1995. Antibiosis and antixenosis of Chrysanthemum cultivars to the aphid Aphis gossypii. Entomologia Experimentalis et Applicata 77:307-314. Tamo M., Baumgartner, J. and Arodokoun, D.Y. 1993. The spatio-temporal distribution of Megalurothrips sjostedti (Trybom) (Thysa-noptera, Thripidae) life stages on cowpea, and development of sampling plans. Bulletin de Societe Entomologique Suisse 66:15-34. Teulon, D.A.J. and Penman, D.R. 1991. Effect of temperature and diet on oviposition rate and development time of the New Zealand flower thrips, Thrips obscuratus. Entomologia Experimentalis et Applicata 60:143-155. Teulon, D.A.J., Penman, D.R. and Ramakers, P.M.J. 1993. Volatile chemicals for thrips (Thysanoptera: Thripidae) host finding and applications for thrips management. Journal of Economic Entomology 36:1405-1415. Trichilo, P.J. and Leigh, T.F. 1988. Influence of resource quality on the reproductive fitness of flower thrips (Thysanoptera: Thripidae). Annals of the Entomological Society of America 81:64-70. Van Emden, H.F. 1987. Cultural methods: the plant. In: Integrated Pest Management. Burn, A.J., Coaker, T.H. and Jepson, P.C. (Eds.), pp.27-68. Academic Press, London. Visser, J.H. 1986. Host odour perception in phytophagous insects. Annual Review of Entomology 31:121-144. Wahl, O. and Ulm, K. 1983. Influence of pollen feeding and physiological condition on pesticide sensitivity of the honey bee Apis mellifera carnica. Oecologia 59:106-128.
Copyright 1998, African Crop Science Society The following images related to this document are available:Line drawing images[cs98006d.gif] [cs98006f.gif] [cs98006c.gif] [cs98006e.gif] [cs98006g.gif] [cs98006a.gif] [cs98006b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
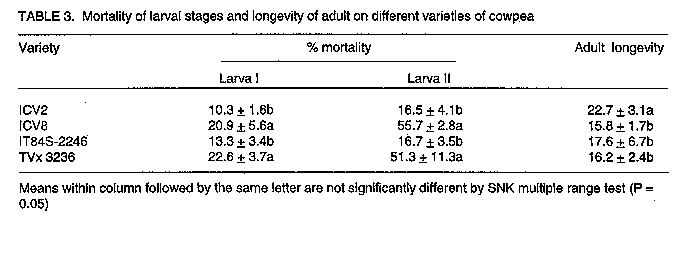{kind=link}
{kind=link}
{kind=link}
{kind=link}