 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 6. No. 2, pp. 109-118, 1998 PERFORMANCE ET STABILITÉ DE RENDEMENT DES GÉNOTYPES DE PATATE DOUCE DANS DIVERS ENVIRONNEMENTS À L'ST DU CONGO P. PHEMBA, T. MUTOMBO, N.B. LUTALADIO1 et E. E. CAREY2 INERA, Centre de Recherche de Mulungu, D.S. Bukavu, Rép. Dém. du Congo 1 PRAPACE, P.O. Box 22274, Kampala, Uganda (Reçu le 23 décembre 1997 et accepté le 20 avril 1998) Code Number:CS98012 RÉSUMÉ Huit génotypes de patate douce d'origine géographique différente étaient évalués durant les saisons culturales des années 1995 et 1996 pour leur performance et leur stabilité de rendement dans sept divers environnements représentatifs du Kivu montagneux à l'est de la République Démocratique du Congo. Les génotypes les plus productifs étaient Yan Shu 1 (16,7 t ha-1), Mulungu 1 (12,2 t ha-1) et Mugande (11,9 t ha-1). Les rendements les plus faibles étaient produits par le clone Nong Da Hong (8,5 t ha-1) et la variété locale Caroline Lee (8,1 t ha-1). Les résultats obtenus de l'analyse de la variance combinée ont montré des différences significatives entre les génotypes, les environnements et les effets d'interaction entre génotype et environnement (G x E) en ce qui concerne le rendement en tubercules. L'analyse de la stabilité du rendement basée sur le coefficient de régression a permis d'identifier deux génotypes stables et à haut rendement à travers les sept environnements. Ces génotypes, Yan Shu 1 et Mulungu 1, ont été ainsi sélectionnés par les chercheurs pour diffusion auprès des producteurs de patate douce à l'est du Congo. Ils pourront aussi être utilisés comme parents dans un programme d'amélioration de la patate douce au Congo. Le comportement des génotypes à travers les altitudes a confirmé que la patate douce est une culture des basses et moyennes altitudes dans les zones à climat chaud. Elle a donné, à l'est du Congo, un rendement acceptable jusqu' à 1.560 m d'altitude au-dessus de laquelle les rendements baissent graduellement pour la plupart des clones à cause des faibles températures entraînant un climat frais d'altitude. Mots Clés: Adaptabilité des génotypes, effets d'environnements, patate douce, stabilité de rendements ABSTRACT Eight sweetpotato genotypes of different geographic origins were evaluated during the 1995 and 1996 growing seasons to determine their performance and stability over seven representative environments in the highlands of Kivu in eastern Congo. Results of combined analysis of variance showed significant differences between genotypes and environments, and significant effects of genotype by environment (G x E) interactions. The highest yielding genotypes were Yan Shu 1 (16.7 t ha-1), Mulungu 1 (12.2 t ha-1) and Mugande (11.9 t ha-1) while the lowest yielding were Nong Da Hong (8.5 t ha-1) and the local Caroline Lee (8.1 t ha-1). Stability analysis of storage root yield, based on the regression coefficient, allowed the identification of high and stable yielding genotypes. Among the clones evaluated, Yan shu 1 and Mulungu 1 were the most stable and high yielding genotypes across all the seven environments. They were retained for release to sweetpotato producers in eastern Congo and would be used as parents in sweetpotato breeding programme in Congo. The response of genotypes of different origins to altitudes showed that sweetpotato is a low and mid-elevation crop of warm climate. It can be grown in eastern Congo to give appreciable yields in altitudes of up to 1,560 m above which yield of most clones gradually decreases due to cooler temperature at higher altitudes. Key Words: Genotype adaptability, effects of environments, yield stability, sweetpotato INTRODUCTION La patate douce (Ipomoea batatas) est une culture importante dans l'alimentation des populations vivant en altitude à l'est de la République Démocratique du Congo et au nord-ouest du lac Victoria en Afrique Orientale (Ewell et Mutuura, 1994). La consommation par habitant est d'environ 84 kg/an (CIP, 1995). Parmi les plantes à racines et à tubercules, la patate douce occupe la deuxième place après le manioc en ce qui concerne la production et la quantité consommée (FAO, 1989; Horton, 1988). Elle est cultivée sur une gamme d'environnements à variations édaphiques et microclimatiques plus ou moins grandes, et à des altitudes allant jusqu' à 2.000 m. Cette région d'altitude à l'est du Congo, couramment appelée le Kivu montagneux, est la principale zone de production de la patate douce en République Démocratique du Congo (Kadiebwe, 1989). Elle couvre une superficie d'environ 87.600 km2 et s'étend sur les altitudes de 800 à plus de 2.600 mètres. Elle est caractérisée par une variabilité écologique observée sur des distances relativement courtes. La relance de la recherche sur l'amélioration de la patate douce dans les pays membres du réseau PRAPACE (Programme Régional d'Amélioration des Patates en Afrique Centrale et de l'Est) est tout à fait récente. Elle s'est consolidée dans une approche régionale en 1992, quand le PRAPACE qui est associé au Centre International de la Pomme de terre (CIP) a intégré la patate douce dans ses activités. Au Congo, la recherche sur la patate douce s'effectue au sein du Programme National de Recherches sur les Tubercules (PNRT) retenu comme l'un des programmes prioritaires de l'Institut National d'Etudes et de Recherches Agronomiques (INERA) au Congo. Le PNRT a pour objectif principal le développement des variétés à rendements élevés, résistantes aux maladies et aux insectes d'intérét économique et adaptées aux divers environnements du Congo. Le germoplasme évalué par le PNRT est d'origine diverse. Il s'agit notamment de cultivars locaux, de variétés en diffusion dans les pays membres du réseau PRAPACE reçues sous forme des boutures indemnes de maladies ainsi que des sélections de la Chine, du Japon, des USA et du Nigéria conservées au CIP, Lima et obtenues en culture in vitro. Le schéma de sélection en cinq saisons utilisé au centre de recherche de l'INERA-Mulungu, commence par l'installation des parcelles d'introduction à la première saison, suivie à la deuxième saison par une évaluation préliminaire, puis à la troisième saison par un essai de rendement préliminaire. Les essais de rendement avancé ont lieu à la quatrième saison et les essais uniformes et multilocaux à la cinquième saison. Beaucoup de génotypes de patate douce évalués par le PNRT sont ainsi déjà arrivés au stade de pré-diffusion. Leur rendement potentiel était évalué en station à l'INERA-Mulungu et sur quelques sites multilocaux, mais leur performance dans différents environnements de l'est du pays n'est pas encore connu. Par conséquent, une série d'essais sur plusieurs et diverses zones agroécologiques avec différents systèmes de production était considérée comme une étape essentielle au développement des cultivars. Cette étape permettra d'identifier les génotypes adaptés aux environnements spécifiques et ceux adaptés à tous les environnements (Allard et Bradshaw, 1964; Kang et Gorman, 1989). Elle fournira ainsi les recommandations sur l'environnement approprié pour l'expression de la potentialité de chaque génotype. Finlay et Wilkinson (1963) ont développé une méthode permettant d'estimer l'adaptabilité des cultivars dans différents environnements, à l'aide de l'analyse de la variance et des coefficients de régression. Eberhart et Russel (1966), Shukla (1972) ainsi que Francis et Kannenberg (1978) ont utilisé des méthodes similaires en définissant d'autres paramètres de stabilité tels que la déviation de la régression, la stabilité de la variance et le groupage des génotypes par classe sur base de leurs rendements moyens et de leur coefficient de variation. Ces méthodes, en plus de celle évaluée par Bacusmo et al. (1988) permettaient d'indiquer qu'un clone se comportait relativement bien dans un bon (b>1,0) ou mauvais (b<1,0) environnement ou présentait un comportement moyen (b=1,0) à travers les différents environnements. Elles facilitaient, en plus, le groupage de plusieurs génotypes selon leurs origines et la comparaison de leur performance et stabilité de rendement dans différents environnements (Francis et Kannenberg, 1978). La présente étude a pour objet d'évaluer la performance et l'adaptabilité des clones de patate douce d'origines différentes et de déterminer la stabilité des génotypes à travers les divers environnements d'altitude à l'est du Congo afin de recommander les génotypes les plus performants pour les environnements appropriés. MATÉRIEL ET MÉTHODES Huit génotypes de sources différentes étaient utilisés pour cette étude. Ces génotypes étaient préalablement évalués pour leur rendement au Centre de Recherches de l'INERA-Mulungu à l'est du Congo. Ils étaient choisis sur la base de leur rendement et de leur précocité relative. Les clones Benikomachi (CIP-440115), White Delite (CIP-440106), Xushu 18 (CIP-440025), Yan Shu 1 (CIP-440024) et Nong Da Hong (CIP-440030) étaient obtenus du CIP sous forme de vitroplants. La variété Mugande (CIP-440163) sélectionnée par l'ISAR au Rwanda était obtenue sous forme de boutures indemnes tandis que la variété Mulungu 1 était sélectionnée par le PNRT à partir des cultivars locaux évalués à Mulungu. La variété Caroline Lee, couramment cultivée par les populations locales du Kivu, était utilisée comme témoin local. Les clones étaient évalués sur sept environnements représentatifs des grandes zones de production de la patate douce des hautes terres du Kivu à l'est du Congo. Ces sept sites choisis avec le concours des paysans étaient différents au point de vue de leur altitude, du type de sol, de la végétation, du précédent cultural et du type de pluviosité (Tableau 1). TABLEAU 1. Attributs caracteristiques des 7 environnements a l'est du Congo ou la patate douce etait cultivee en 1995 et 1996
L'essai était conduit sur les sept sites durant les saisons culturales de 1995 et 1996, dans un dispositif expérimental en blocs complètement randomisés en quatre répétitions. Un écartement de 1,00 m entre les billons et de 0,30 m sur les billons était utilisé dans des parcelles élémentaires comprenant 4 billons de 6 mètres chacun, soit une superficie unitaire de 24 m2. Des boutures apicales de patate douce, d'environ 30 cm de longueur, étaient choisies pour la plantation. L'essai était planté à la méme période, en mars et novembre de chaque année, dans les différents sites. Une ligne de bordure de patate douce était plantée autour de l'essai. Pendant la croissance des plantes, l'essai n'a reçu ni fertilisation ni traitement phytosanitaire, à l'exception du sarclage. La récolte a eu lieu après quatre mois de plantation. Elle a été effectuée en présence et avec la collaboration des paysans. Ceux-ci ont donné leur appréciation sur le nombre, la forme et la couleur de tubercules de chaque clone. La récolte était faite manuellement; les tiges de la patate douce étaient coupées et pésées. A l'aide d'une houe, les tubercules étaient déterrés systématiquement, classés puis séparés visuellement en tubercules larges ou commercialisables ayant un diamètre d'au moins 4 cm et en petits tubercules non commercialisables. Les données étaient récoltées sur deux billons à l'intérieur de la parcelle unitaire pour déterminer le nombre de tubercules, le rendement total et le rendement en tubercules commercialisables. L'indice de récolte était déterminé par le pourcentage du poids de tubercules par rapport au poids total de la plante. Les données sur les dégëts causés par les ravageurs (Cylas sp.), la réaction aux viroses ainsi que sur la préférence selon le goût et la couleur de tubercules étaient également enregistrées. Les résultats obtenus étaient analysés en calculant la variance et les erreurs standard de toutes les variables mesurées. Une analyse de la variance combinée était faite pour le rendement total de tubercules en utilisant le programme MSTAT-C (Anonymous, 1990) afin de tester l'effet de l'année, de l'environnement et du génotype, et les effets d'interactions. Les tests de Fisher étaient utilisés pour établir les niveaux de signification. Une analyse de la stabilité de rendement en tubercules pour les 8 génotypes à travers tous les environnements était faite en se basant sur les méthodes développées par Eberhart et Russel (1966), Francis et Kannenberg (1978) et Finlay et Wilkinson (1963). La méthode d'Eberhart et Russel (1966) utilise le modèle de régression Yij = Ui Bi Ij + Sij, où Yij est la moyenne de ith génotype à jth environnement, U la moyenne du ith génotype à travers tous les environnements, Bi le coefficient de régression qui mesure la réponse de ith génotype à la variation des environnements, Ij l'indice de l'environnement et Sij la déviation de la régression de ith génotype à jth environnement. Les coefficients de régression (b) étaient calculés pour chaque variété en régressant la moyenne de chaque environnement sur la moyenne totale de tous les génotypes dans chaque environnement en utilisant la formule b = SYij Ij/SI2j. Selon cette méthode, un clone stable est celui qui a un coefficient de régression (b = 1,0) et une très faible déviation de la régression. La méthode de Francis et Kannenberg (1978) groupe les génotypes en 4 classes sur la base de leurs rendements moyens et de leurs coefficients de variation. La classe 1 comprend les génotypes ayant des rendements moyens élevés et des faibles coefficients de variation. La classe 2 est constituée de génotypes ayant des rendements moyens élevés et des coefficients de variation élevés. La classe 3 regroupe les génotypes à faibles rendements moyens et à faibles coefficients de variation. La classe 4 comprend les génotypes ayant des faibles rendements moyens et des coefficients de variation élevés. La méthode de Finlay et Wilkinson (1963) met en relation graphique le coefficient de régression de chaque génotype et son rendement moyen transformé en logarithme pour induire la linéarité. Les coefficients de régression étaient testés pour leur signification par le test-t de Student, les déviations de régressions et les coefficients de détermination étaient également calculés. RÉSULTATS ET DISCUSSION Des différences significatives étaient observées dans les sept environnements pour le rendement en tubercules des huit génotypes de patate douce. Les génotypes Yan Shu 1, Mulungu 1 et Mugande ont donné des rendements élevés à travers les différents environnements tandis que les génotypes Nong Da Hong et Caroline Lee avaient des faibles rendements. D'autres indicateurs d'effets d'environnement étaient mis en évidence par les différences significatives en nombre de tubercules et en indice de récolte (Tableau 2). TABLEAU 2. Rendement et composants du rendement de 8 genotypes de patate douce dans 7 environnements a l'est du Congo (1995 et 1996)
L'analyse de la variance combinée du rendement en tubercules a montré des effets significatifs (Tableau 3) qui indiquent que les génotypes avaient un comportement différent dans chaque environnement (Allard et Bradshaw, 1964; Kang et Gorman, 1989). Les interactions du premier et du second ordre étaient significatives indiquant que chaque environnement de chaque année était différent (Tableau 3). TABLEAU 3. Analyse combinee de la variance de 8 genotypes de patate douce dans 7 environnements a l'est du Congo (1995 et 1996)
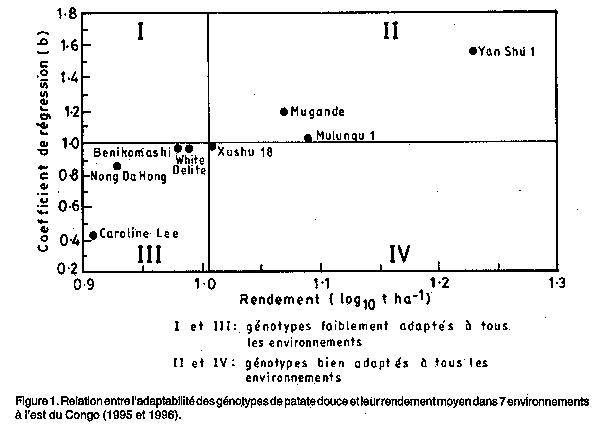
*, ** significatif a 5% et 1% de niveau de probabilite, respectivement TABLEAU 4. Rendement moyen et parametres de stabilite de 8 genotypes de patate douce dans 7 environnements a rest du Congo (1995 et 1996)
b: pente de regression Les paramètres de stabilité de rendement en tubercules varient de 0,43 à 1,56 (Tableau 4). Ces paramètres montrent que les huits génotypes réagissent différemment aux effets d'environnement. Les génotypes Mulungu 1, Xushu 18, White Delite et Benikomachi ont des pentes de régression proches de l'unité (b = 1,0) suggérant qu'ils sont relativement plus stables que les autres génotypes (Tableau 4). Selon Eberhart et Russel (1966), la moyenne des quarrés de la déviation de régression (S2d) est la vraie mesure de stabilité. Un génotype stable serait donc celui qui aurait un rendement plus élevé que la moyenne des rendements, un coefficient de régression b = 1,0 et une moyenne des quarrés de la déviation de régression, S2d, proche de 0. Suivant cette définition, seul le génotype Mulungu 1 peut étre considéré comme étant très stable à travers tous les environnements de cette étude (Tableau 4). Les autres génotypes ne peuvent plus étre stables quant les environnements deviennent défavorables, comme l'a suggéré Finlay et Wilkinson (1963). Les résultats obtenus montrent aussi que la corrélation entre le rendement moyen en tubercules et la moyenne des carrés de la déviation est significative et positive (Tableau 4). Ceci indique que les génotypes à haut rendement, à l'exception de Mulungu 1, ont des valeurs de déviation élevées, qui ne sont pas toujours désirables (Abebe et al, 1995), ainsi que des valeurs élevées du coefficient de détermination de régression. L'analyse de la régression a une valeur prédictive qui permet ainsi l'estimation de la stabilité des génotypes (Assefa et al., 1995). L'interprétation de l'adaptabilité des génotypes est utilisée pour évaluer la relation entre les génotypes et leurs origines. La figure 1 est obtenue par la relation entre les coefficients de régression des moyennes de rendements des génotypes et le logarithme de leurs rendements moyens. Elle montre que les génotypes d'origine asiatique tels que Xushu 18, Benikomachi et Nong Da Hong sont, à une exception près, groupés. Ces génotypes, à l'exception de Yan Shu 1 mais ensemble avec White Delite d'origine américaine et la variété locale Caroline Lee, ont des valeurs faibles de coefficient de régression et des rendements plus faibles que la moyenne. Ils ont donc tendance à étre mal adaptés à tous les environnements. Par contre, les génotypes Yan Shu 1, Mugande et Mulungu 1, qui ont des rendements plus élevés que la moyenne et des coefficients de régression supérieurs à l'unité, sont bien adaptés à tous les environnements (Fig. 1). Le groupage des génotypes par la méthode de Francis et Kannenberg (1978) a permis d'identifier les génotypes Yan Shu 1 (CIP-440024) d'origine chinoise et Mulungu 1 (sélection locale) comme étant stables et très performants à cause de leurs rendements élevés et de leurs faibles variations de coefficient de variation (Fig. 2). Le clone Mugande d'origine rwandaise a été jugé instable parce que tout en ayant un rendement élevé, il présente une grande variation du coefficient de variation. Les clones Xushu 18, Nong Da Hong et Caroline Lee, respectivement d'origine chinoise et locale, sont cotés très instables parce qu'ils ont des rendements plus faible que la moyenne et des coefficients de variations très élevés. Enfin, les clones Benikomachi et White Delite, respectivement d'origine japonaise et américaine, sont assez stables bien que leur rendement soient plus faible que la moyenne (Fig. 2). Figure 1. Relation entre l'adaptabilité des génotypes de papate douce et leur rendement moyen dans 7 environnements à l'est du Congo. Figure 2. Relation entre le rendement moyen de 8 papate douce et leur stabilité phénotypique dans 7 environnements à l'est du Congo (1995 et 1996). Figure 3. Rendement moyen des groupes d'origines des génotypes de papate douce dans sept environnements à l'est du Congo (1995 et 1996). La réponse des génotypes vis-à-vis des altitudes est présentée à la Fig. 3 qui montre clairement que la patate douce à l'est du Congo est une plante de moyenne altitude, jusqu'à 1.560 m d'altitude environ, au-dessus de laquelle les rendements sont en général graduellement réduits. Les génotypes d'origines différentes ont montré, à l'exception de la variété locale, un comportement similaire selon les altitudes. Cependant, il y avait une réduction brusque du rendement à 1.370 m d'altitude pour tous les génotypes à l'exception de la variété locale, probablement à cause de l'état marécageux des sols de ce site (Ngongo, communication personnelle, INERA, mai 1997). Les clones exotiques dont Yan Shu 1 semblent s'adapter au climat chaud tandis que Mulungu 1 et Mugande semblent se comporter assez bien en milieu marécageaux. En outre, les génotypes d'origines diverses ne se sont pas bien comportés à partir de 1.920 m d'altitude à cause probablement des basses températures et des faibles intensités de lumière entraönant un climat relativement frais qui ne favorisait pas le développement des tubercules. Suivant la performance et l'analyse de stabilité décrite par Eberhart et Russel (1966), les génotypes Yan Shu 1 et Mulungu 1 sont les plus productifs et stables à tous les environnements considerés. Ils pourront ainsi étre cultivés et diffusés à travers les différents environnements du Kivu montagneux et étre utilisés comme parents dans un futur programme d'amélioration de la patate douce à l'est du Congo. REMERCIEMENTS La contribution des agronomes techniciens du PNRT et des paysans qui ont collaboré à cette étude est hautement appréciée. Les auteurs remercient le Centre de Recherche de l'INERA-Mulungu pour le support logistique et administratif. Ils sont très reconnaissants au CIP, au PRAPACE et au CRDI (Centre de Recherche pour le Développement International) pour avoir, respectivement, fourni le matériel végétal éxotique, l'appui scientifique et le soutien financier sans lesquels ce travail n'aurait pas pu se réaliser. RÉFÉRENCES Abebe Menkir, Yilma Kabede and Berhane Gebrekidan. 1984. Genotype-environment interaction and yield stability in sorghum of intermediate maturity. Ethiopian Journal of Agricultural Science 6:1-11. Allard, R.W. and Bradshaw, A.D. 1964. Implications of genotype-environment interactions. Crop Science 4:503-507. Anonymous. 1990. MSTAT-C: A microcomputer program for the design, management and analysis of agronomic research experiments. Michigan State University, East Lansing, MI. Assefa Solomon, Geleta Bekele and Tanner, D.G. 1995. Yield stability analysis of nine spring bread (Wheat) genotypes on the Central Highlands of Ethiopia. African Crop Science Journal 3:35-40. Bacusmo, J.L., Collins W.W. and Jones, A. 1988. Comparison of methods of determining stability and adaptation of sweetpotato. Theor. Appl. Genet. 75:492-497. CIP. 1995. La pomme de terre en chiffres: production, utilisation, consommation et commerce. Un abrégé d'information-clefs. Eberhart, S.A and Russel, W.L. 1966. Stability parameters for comparing varieties. Crop Science 6:36-40. Ewell, P.T. and Mutuura, J. 1994. Sweetpotato in the food systems of eastern and southern Africa. In Tropical root crops in a developing economy. Ofori, F. and Hahn, S.K. (Ed.) Proceedings of the ninth symposium of the International Society for Tropical Root Crops, 20-26 October 1991, Accra, Ghana. pp. 405-412. FAO. 1989. FAO Production Yearbook 1988. FAO, Rome. Finlay, K.W. and Wilkinson, G.N. 1963. The analysis of adaptation in a plant breeding programme. Australian Journal of Agricultural Research 14:742-754. Francis, T.R. and Kannenberg, L.W. 1978. Yield stability studies in short season maize. I. A descriptive method for grouping genotypes. Canadian Journal of Plant Science 58:1029-1034. Horton, D. 1988. Underground crops: Long-term trends in production of root and tubers. Winrock International Institute for Agricultural Development (USA) Morrilton. pp 130 Kadiebwe, N. 1989. Situation de la culture de la patate douce en Ituri au Zaòre. Bulletin d'Information de l'IRAZ No. 2:8-19. Kang, M.S. and Gorman, D.P. 1989. Genotype x environment interaction in maize. Agronomy Journal 81:662-664. Shukla, G.K. 1972. Some statistical aspects of partitioning genotype-environment components of variability. Heredity 29:237-245. Copyright 1998, African Crop Science Society The following images related to this document are available:Line drawing images[cs98012a.gif] [cs98012c.gif] [cs98012b.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}