 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
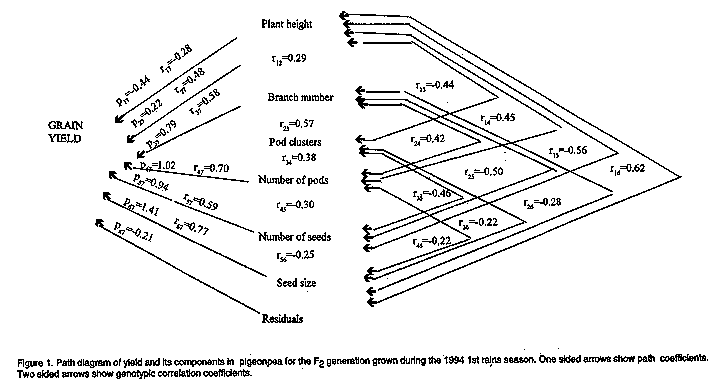
African Crop Science Journal, Vol. 6. No. 2, pp. 143-148, 1998 PATH COEFFICIENT ANALYSIS OF YIELD AND ITS COMPONENTS IN PIGEONPEA M.S. Musaana and M.S. Nahdy Kawanda Agricultural Research Institute (KARI), P.O. Box 7065, Kampala, Uganda (Received 16 May, 1997; accepted 5 March, 1998) Code Number:CS98016 ABSTRACT Eight pigeonpea (Cajanus cajan (L.) Millsp.) lines previously inbred for two seasons were crossed in a diallel system. The parents, F1 and F2 populations were used to record yield and some of the yield components. Phenotypic and genotypic correlations were calculated from the data, and later a path coefficient analysis carried out using the genotypic correlations. Both the phenotypic and genotypic correlations showed that plant height, pods/plants, seeds/pods, seed weight and primary branch number were the primary components of yield in pigeonpea. A path coefficient analysis, however, indicated that pod clusters/plant, pods/plant, seeds/pod and seed weight were the main yield components having maximum direct effects on yield. Key Words: Breeding, correlations, diallel crossing, genotypic, phenotypic, reciprocal regressions RÉSUMÉ Huit lignées de pois (Cajanus cajan (L.) Millsp.) préalablement consanguins pendant deux saisons ont été croisés selon un système diallèle. Les parents, les populations F1 et F2, ont été utilisés afin d'enregistrer le rendement et d'autres paramètres du rendement. Les correlations phénotypiques et génotypiques ont été calculées à partir des données obtenues et une analyse du coefficient de passage a été faite en utilisant les correlations génotypiques. Les deux correlations, phénotypiques et génotypiques, ont montré que la hauteur de la plante, les rapports gousses/plantes, graines/gousses, le poids de graines et le nombre de branches primaires étaient les principaux composants du rendement de pois cajan. Une analyse du coefficient de passage a cependant montré que les rapports bouquets de gousses/plantes, gousses/plantes graines/goussess et le poids de graines étaient les principaux composants du rendement ayant des effets directs maximum sur le rendement. Mots Clés: Croisement, correlations, croisement diallèle, génotypique, phénotypique, régressions réciproques INTRODUCTION Pigeonpea (Cajanus cajan (L.) Millsp.) is an important grain legume in Uganda. It is consumed as dry seed and green peas for vegetables. It also offers an excellent livestock feed both as dry seed and as forage. The seeds contain 18-29% protein on dry weight basis which is about three times the value found in cereals. The protein is also of excellent quality, being high in lysine (Oland and Stabursuik, 1970; Rachie and Wurster, 1971). The crop is, therefore, an important complement to cereals, root crops and bananas which are the major constituents of the diet of the people of Uganda. In view of the importance of the crop and the low average yields in Uganda, an improvement programme was initiated at Kawanda Agricultural Research Institute (KARI) in 1992 to develop high yielding varieties. Such varieties had to be developed using the current low yielding varieties as some of the parents by using proper breeding procedures. However, breeding new lines could only be undertaken after the effects of individual components on yield had been determined. A path-coefficient analysis of yield and its components was, therefore, carried out on the data collected from a diallel crossing system. Path coefficient analysis for yield and its components by previous workers had shown that pod clusters per plant was the major determinant of yield in pigeonpea (Singh and Malhotra, 1973; Veeraswamy et al., 1973). Similar results have been reported on cowpea by Musaana (1979) but S'Ozdemir (1996) reported that clusters per plant, pods per plant and seed weight contributed more to grain yield in chickpea. Pigeonpeas as a crop have yields that range between + 1200kg ha-1 to + 100 kg ha-1 depending on the cultivar. Pods are borne at intervals on clusters that may bear one to 10 pods per cluster. The pods themselves vary from < 50 pods on poor yielding annuals to > 200 pods on perennial or high yielding plants. The number of seeds per pod usually varies between 5 and 10. MATERIALS AND METHODS During the first rains in the 1993 season, a complete diallel cross was made using eight lines that were inbred for two generations. The lines were selected for wide differences in yield and yield components. Half of the F1 seed was grown in isolation to get F2 seed. All possible F1's (including reciprocals), F2's and parents were grown in the field in a randomised complete block design with 4 replicates. Each plot measured 4 x 4m. The spacing was 1m between rows and 0.5m within rows. At maturity plant height and width, primary branch number, pod clusters and pods per plants, seeds per pod and seed weight were recorded on fifty randomly selected plants per plot. Phenotypic correlation coefficient for the F1 and F2 generations were determined by the variance/covariance method (Falconer, 1960) and genotypic correlation were calculated for the F2 generation using the method suggested by Fisher (1963). By using the genotypic correction coefficients a path coefficient analysis was carried out using the method suggested by Dewey and Lu (1959). The path coefficients in this paper were obtained through simultaneous solution of the equations which express all possible two-way relationships among variables (i.e., relationships between collerations and path coefficients). The path coefficients are thus direct effects expressed as standardised partial regressions coefficients which can be greater than one and are expressed in standard deviation units. RESULTS AND DISCUSSIONS Phenotypic and genotypic correlations for the F2 populations of the diallel cross for yield and yield components are presented in Table 1. The phenotypic correlations were lower than the corresponding genotypic correlations. However, both phenotypic and genotypic correlations were close to each other in each instance. This may have been due to the large number of plants (50) used to take observations per plot which tended to reduce the error environmental variance to minor proportions. If the environmental variance was reduced to zero, the phenotypic and genotypic correlations would have been identical as was pointed out by Dewey and Lu (1959). Since the two types of correlations were similar, reference will be made only to genotypic correlations to avoid repetitions. TABLE 1. Phenotypic and genotypic correlations between yield with its components in the F1 generation of the diallel cross
*, ** Significant at 5% and 1% levels, respectively It was only plant height that showed a negative correlation with yield. Plant height and seeds per pod were also negatively and significantly (P<0.01) correlated. Seeds per pod and seed weight were negatively correlated with branch number, cluster number and pods/plants. Seeds/pod was also negatively correlated with seed weight. However, plant height and seed weight (100 seed weight) were positively and significantly correlated (r = 0.62, P<0.01). This may have been due to the large leaf surface which photo-synthesised higher quantities of carbohydrates that were directed to seed formation. This combined with the fact that larger plants produced fewer seeds/pod, gave larger plants which is an advantage over smaller plants with respect to seed weight. Pathnaswamy et al. (1973) also reported that the seeds of perennials were larger and heavier that those of annuals. Many of the characters were negatively or positively correlated due to mutual association with other characters. Results of path coefficient analysis to separate out the direct and indirect causes of association are presented in Table 2 and Figure 1. Yield vs. plant height. The correlation between grain yield and plant height was negative and non-significant (r = -0.28). The direct effect of plant height on yield was high and negative (-0.44). The indirect effect of plant height on seeds/pod was high and negative (-0.56) and the indirect influences of plant height on yield though branch number (0.29) and seed weight (0.25) were both low and positive. The net effect in the system of opposing influences was that two large negative effects counterbalanced two positive ones making the overall correlation between plant height and grain yield low and negative. The results indicated that plant height was not an important yield component. Yield vs. number of primary branches. The total correlation between yield and branch number was high positive and significant (r = 0.48,P<0.05). This resulted from the high positive, indirect influences of branch number on grain yield via cluster number and pods per plant of 0.46 and 0.42, respectively. But the direct influence of branch number on grain yield was low (0.22) due to the high negative indirect influence of branch number on grain yield via seeds/pod and seed size. This was because highly branching plants matured late in the season when pest populations had built up (Cruz, 1975). The results show that branch number was not an important yield component. Yield vs. clusters per plant. The total correlation of cluster number with yield was high and positive, (r = 0.58, P<0.01). The direct effect of clusters per plant on yield was very high (0.79) and positive. Clusters per plant also registered moderately high indirect and positive effects on yield via branch number (0.46) and pods per plant (0.38). The negative, indirect influences of clusters per plant were registered though seed size (-0.20) and seeds/pod (-0.46). The results indicated that pod clusters/plant were important yield components. Similar views were expressed by Singh and Malhotra (1973). Yield vs. pods per plant. The correlation between pods per plant and grain yield was high, positive and significant (r = 0.70, P<0.01). The direct effect of pods per plant on yield was also very high and positive (1.02). A positive, indirect influence of pods per plant on yield was registered via branch number (0.42). Negative, indirect influences of pods/plant on yield were registered via seeds per pod (-0.30) and seed size (-0.33). The high and positive direct influence of pods/plant on yield indicated that this was an important yield component. Veeraswamy et al. (1973) also reported that pods/plant was an important yield component in pigeonpea. Yield vs. seeds per pod. The correlation between seeds per pod and yield was high and positive (r = 0.59, P<0.01). The direct effect of seeds per pod on yield was also high (r = 0.94). The negative indirect effects of seeds per pod on grain yield were low and unimportant. The net effect was that seeds/pod was an important yield component. Contrary results were reported by Singh and Malhotra (1973) and Veeraswamy et al. (1973) that only pod clusters/plant and pods/plant were important yield components in crop. Yield vs. seed weight. The correlation between yield with seed size was high, positive and significant (r = 0.77, P<0.01). The direct effect of seed size on grain yield was very high (1.41), while indirect effects of seed size on yield via branch number (-0.28), clusters per plant (-0.22), pods per plant (-0.22) and seeds per pod (-0.25) were negative, but unimportant. Seed size was, therefore, an important yield component. The residuals were negative and non-significant indicating that there was no need to include more yield components like plant width in the path-coefficient analysis. TABLE 2. Partition of genotypic correlation into direct and indirect effects by path coefficient analysis
*,** Significant at 5% and 1% levels, respectively Figure 1. Path diagram of yield and its components in pigeonpea for the F2 generation grown during the 1994 1st rains season. CONCLUSION Significant (P<0.05) and positive phenotypic correlations were found for grain yield with plant height, pods/plant, seeds/pod, seed size and number of primary branches. The relationship between plant height and grain yield was negative though non-significant. These findings tentatively suggested that these characters were the primary components of yield in pigeonpea. A path coefficient analysis, however, showed that clusters per plant, pods/plant, seeds/pod and seed size were the main yield components having maximum direct effects on yield, as previously reported (Singh and Malhotra, 1973; Veeraswamy et al., 1973). Therefore, selection for a combination of these characters would be effective in increasing yield in pigeonpea. Since heavy seeds were fewer in a pod, medium weight seed types on plants having many pods and clusters per plant would probably give the best yields. REFERENCES Cruz, C. 1975. Observation of pod borer oviposition and infestation of pigeon pea varieties. Journal Agriculture University P.R. 59:63-68. Dewey, D.R. and Lu, K.H. 1959. A path coefficient analysis of components of crested wheat grass seed production. Agronomy Journal 51:515-518. Falconer, D.S. 1960. Introduction to Quantitative Genetics. The Ronald Press Co., New York. 186pp. Fisher, R.A. 1963. Statistical Methods for Research Workers. 13th Edn. Oliver and Boyd, London. Musaana, M.S. 1979. Heritability of and Environmental Influence on Major Yield and Agronomic Characters of Pigeon peas. MSc. Thesis, Makerere University, Kampala. Oland, K. and Stabursvik, A. 1970. Requirements of proteins and amino acid availability in Uganda. Memeograph, 10pp. Pathnaswamy, R., Veeraswamy, R., Regupathy, A. and Palaniswamy, G.A. 1973. Studies on genetic variability of certain quantitative characters in red gram (Cajanus cajan (L.)Millsp.) as a horticultural crop in East Africa. First East African Horticultural Symposium held in Kampala, Uganda, 1970. Singh,K.B. and Malhotra,R.S. 1973. Yield components in pigeon pea (Cajanus cajan (L.) Millsp.). Madras Agricultural Journal 60:364-366. S'Ozdemir. 1996. Path coefficient analysis for yield and its components in chick pea. International Chickpea and Pigeonpea Newsletter 3:19-21. Veeraswamy, R., Rangaswamy, P., Fazlullah Khau, A.K. and Muhammed Shereef. 1973. Heterosis in Cajanus cajan (L.) Millsp. Madras Agricultural Journal 60:1317-1319. Copyright 1998, African Crop Science Society The following images related to this document are available:Line drawing images[cs98016a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}