 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
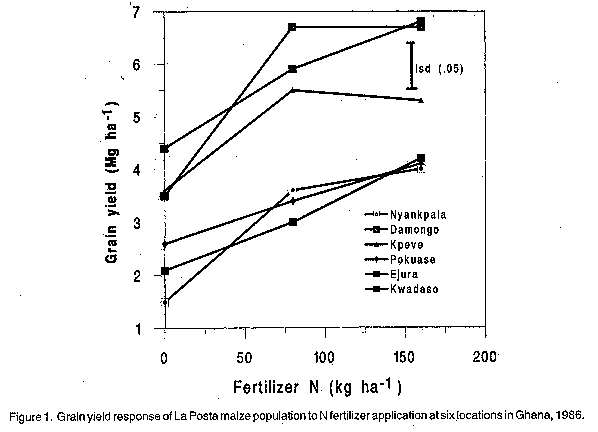
African Crop Science Journal, Vol. 6. No. 3, pp. 241-248, PROGRESS FROM SELECTION IN LA POSTA MAIZE POPULATION EVALUATED UNDER THREE NITROGEN FERTILIZER LEVELS P.Y.K. SALLAH, N.J. EHLKE1 and J.L. GEADELMANN2 Crops Research Institute, CSIR, P.O. Box 3785, Kumasi, Ghana (Received 15 January, 1998; accepted 15 June, 1998) Code Number:CS98026 ABSTRACT Nitrogen (N) is an important nutrient whose application directly influences the yield of maize (Zea mays L.) and other non-leguminous crop plants. In 1986, cycles 0, 2, 4, and 6 of CIMMYT Population 43, improved using full-sib family selection for grain yield and other traits, were evaluated at 0, 80, and 160 kg N ha-1 at six locations in Ghana, West Africa. Grain yields across environments and cycles averaged 3.0, 4.7 and 5.2 Mg ha-1 at 0, 80 and 160 kg N ha-1, respectively. Cycle responses showed that grain yield, plant height, lodging, and ear rating (based on size and uniformity of ears) increased, days to mid-silk decreased, and grain moisture and number of ears per plant did not change as fertilizer N was increased. The greatest response to applied N was observed at 80 kg N ha-1 for all traits. Similar N response patterns were observed in all cycles. Least square estimates of progress per cycle of selection for grain yield, days to mid-silk, plant height, and lodging were 1.6, -0.6, -1.5, -1.1% at zero fertilizer N; 2.1, -0.8, -1.5, -3.5% at 80 kg N ha-1 and 1.8, -0.7, -1.4, -2.8% at 160 kg N ha-1, respectively. These results indicate that (i) selection was effective for improving grain yield, days to mid-silk, plant height, and lodging in the population at 80 and 160 kg N ha-1, and (ii) recurrent selection for improved agronomic performance had little effect on the N fertilizer response of the population. Key Words: Genetic gain, nitrogen response, recurrent selection, selection cycles, Zea mays RÉSUMÉ L'azote (N) est un élément nutritif important dont l'application influence directement le rendement du maïs (Zea mays L.) et d'autres cultures non légumineuses. En 1986, les cycles améliorés de 0, 2, 4, et 6 de Population 43 de CIMMYT utilisant èla sélection familiale des vrais parents' pour le rendement en grains étaient évalués à 0, 80 et 160 kg/ha d'azote dans six localités du Ghana, Afrique de l'Ouest. Les rendements en graines à travers les environnements et les cycles étaient en moyenne de 3.0, 4.7 et 5.22 Mg/ha respectivement à 0, 80 et 160 kg/ha d'azote. Les cycles de réponse ont montré que le rendement en grains, la taille de plant, la verse, et le taux d'épi (basé sur la grandeur et l'uniformité des épis) ont augmenté, les jours à la mi-soie ont diminué, l'humidité des grains et le nombre d'épis par plant n'ont pas changé quand la quantité d'azote a été augmentée. La réponse la plus significative à l'application de l'azote a été observée à 80 kg/ha d'azote pour tous les traits. Les tendances similaires des modèles de réponse à l'azote ont été observées dans tous les cycles. Les estimations des progrès de moins carrés par cycle de sélection pour le rendement en grains, le nombre de jours à la mi-soie, la hauteur de plant et la verse étaient 1.6, -0.6, -1.5, -1.1 % à 0 kg/ha d'azote, 2.1, -0.8, -.1.5, -3.5 % à 80 kg/ha de N et 1.8, -0.7, -1.4, -2.8 % à 160 kg/ha d'azote, respectivement. Ces résultats montrent que (i) la sélection était efficace pour améliorer le rendement en grain, le nombre de jours à la moitié de soie, la hauteur de plant, et la verse dans la population à 80 et 160 kg/ha d'azote et (ii) la sélection récurrente pour la performance agronomique améliorée a eu peu d'effet sur la réponse de l'engrais azoté de la population. Mots Clés: gain génétique, réponse à l'azote, sélection récurrente, cycle de sélection, Zea mays INTRODUCTION Maize (Zea mays L.) is an important crop under rain-fed agriculture in the lowland tropics. Most cultivated soils in these regions, particularly the savanna soils are deficient in N (Djokoto and Stephens, 1961; Ahn, 1970; Sanchez, 1976). For this reason, N fertilizers are usually recommended for profitable maize production. Nitrogen supply to plants promotes leaf area development, delays leaf senescence (Novoa and Loomis, 1981; Muchow, 1988a) and increases the photosynthetic capacity of the leaf canopy (Murata, 1969; Lugg and Sinclair, 1981). Increase in the N supply may influence the grain yield of maize through increase in the harvest index (Viets, 1965; Muchow, 1988b), average ear weight, weight of the second ear in two-eared varieties, and number of ears per plant in multiple ear hybrids (Krantz and Chandler, 1954). Nitrogen utilisation (N uptake and translocation to developing kernels) in maize is under polygenic control (Harvey, 1939; Pollmer et al., 1979) and variation exists among maize genotypes for N utilisation (Harvey, 1939; Chevalier and Schrader, 1977; Pollmer et al., 1979). Genotypic variation for N utilisation has permitted selection for this trait (Muruli and Paulsen, 1981; Moll et al., 1987). Recurrent selection programmes traditionally develop maize populations under high levels of N fertilisation. This practice seeks to eliminate N as a variable in order to facilitate selection of superior genotypes (Balko and Russell, 1980; Muruli and Paulsen, 1981). Since genotypic variation for N utilisation exists in maize, the N response of the original population may be altered in successive cycles through recurrent selection for improved agronomic performance (Allan and Darrah, 1978; Muruli and Paulsen, 1981; Kamprath et al., 1982). For example, Kamprath et al. (1982) reported that improved population hybrids produced more grain per unit of applied N than the original population hybrids. Further evidence from the U.S. Corn Belt shows that modern maize cultivars (single cross hybrids) are more efficient in utilisation of fertilizer nutrients than the open-pollinated varieties and four-way hybrids of the 1930's (Castleberry et al., 1984; Carlone and Russell, 1987). However, little information is available for maize adapted to the lowland tropics. This study was undertaken (1) to determine if changes have occurred for response to N fertilizer in a lowland tropical maize population following six cycles of conventional recurrent selection and (2) to estimate genetic gains from full-sib family selection for grain yield and other important traits in the population under the different rates of applied N. MATERIALS AND METHODS The N response of cycles 0, 2, 4 and 6 of La Posta maize population was studied in Ghana, West Africa. La Posta corresponds to CIMMYT Population 43 and is a white dent, late maturing Tuxpeno-related synthetic which produces tall vigorous-growing plants, and is well adapted to the lowland tropics (Pandey et al., 1986; CIMMYT, 1987). Cycles 1 and 2 were developed using full-sib selection in which only among-family selection was done. Cycles 3 through 6 were developed using a modified full-sib selection in which both among- and within-family selection was practised. Both selection schemes involved international progeny testing in Mexico and in at least three other countries in the lowland tropics. Two hundred and fifty full-sib families were evaluated in each test. Based on the data from the tests and from a high plant density stress nursery at CIMMYT, Mexico, approximately 40% superior families were selected, using an independent culling procedure. Grain yield, days to mid-silk, plant and ear heights, lodging, and disease reaction were the major traits considered for improvement. In addition, the C5 and C6 populations were improved for resistance to the maize streak virus at the International Institute of Tropical Agriculture (IITA), Ibadan through a CIMMYT-IITA collaborative research effort. Recombination and formation of full-sibs by reciprocal mating of selected individuals from different full-sib families was done in Mexico. Details of the selection schemes have been well described (Pandey et al., 1986). At CIMMYT, the full-sib progenies were formed and tested under 200 kg N ha-1. The N rates used to evaluate the progenies in the other countries depended on fertilizer recommendations in each country. These rates were generally lower than those used at CIMMYT. For example, 90 kg N ha-1 was applied in the progeny tests conducted in Ghana in 1976 (cycle 2), 1978 (cycle 3) and 1980 (cycle 4). Fresh seeds of the cycles evaluated in this study were kindly provided by CIMMYT. The cycles were grown in Ghana in 1986 at Nyankpala (Guinea savanna, fine sandy-loam Alfisol), Damongo (woodland savanna, fine sandy-loam Alfisol), Pokuase (coastal savanna, fine sandy-loam Dystrochrept), Ejura and Kpeve (forest-savanna transition, fine-coarse sandy-loam, Oxisol), and Kwadaso (forest, coarse sandy-loam Paleustult). The previous crop was cowpea (Vigna unguiculata L. Walp.) at Kpeve, Damongo and Ejura. The fields at Nyankpala, Kwadaso and Pokuase were under fallow since the previous maize crop in 1983. The experimental design was a randomised complete block arranged in a split-plot with four replications at each site. Nitrogen rates were assigned to main plots and selection cycles were the sub-plots. Sub-plots consisted of two 5-m rows spaced 0.8 m apart. Hills were spaced 0.5 m, over-planted and thinned to two plants per hill at the 6-to 8-leaf stage to give a uniform population density of 50,000 plants ha-1. All trials were rain-fed except at Kwadaso where supplemental irrigation was applied. Nitrogen rates of 0, 80, and 160 kg N ha-1 were applied as ammonium sulphate in a split dose at two and four weeks after planting. In each case, the N fertilizer was evenly surface-applied to all maize hills within the plot. Phosphorus was broadcast over all plots at 60 kg P205 ha-1 as triple superphosphate prior to planting. No K was applied because of consistent lack of response of maize to K fertilisation in Ghana (Ahn, 1970; Dennis, 1983). Pre-emergence chemical weed control was practised but supplemental hand-weeding was also done when necessary to keep plots free of weeds. Data were recorded on bordered two-plant hills within each plot on days to mid-silk, plant height, total lodging, ears per plant, ear acceptability, grain moisture, and grain yield. Grain yield was adjusted to 150 g kg-1 moisture. Lodging was visually scored on a 1 to 5 scale, where 1 = all plants erect and 5 = all plants broken below the ear or leaning at an angle greater than 450. Ears were visually scored for acceptability on 1 to 5 scale, where 1 = good and 5 = poor, based on size and uniformity of ears, kernel rows, and damage from ear rots and insects. Plant height was measured in centimetres as the distance between the base of the plant and the point where tassel branching began. Number of ears per plant was calculated as the total number of ears harvested divided by the number of plants harvested. Days to mid-silk was calculated as number of days from planting to when 50 percent of the plants per sub-plot had visible silks. Cycles of selection and N levels were considered fixed effects and environments were considered random effects. Analyses of variance were computed in each environment (location) and combined over environments for all traits (Steel and Torrie, 1980). Response to selection was estimated as the regression coefficient (b-value) of the linear regression of the mean value of trait on number of selection cycles and expressed as a percent of the cycle 0 mean (Hallauer and Miranda, 1981). The variation among cycles was partitioned into sums of squares due to linear regression and deviations from the model and F-tests were used to determine the significance levels of the b-values (Hallauer and Miranda, 1981; Steel and Torrie, 1980). RESULTS AND DISCUSSION The combined-over-environments analyses of variance for five traits measured are presented in Table 1. The effects due to environments (E), N, and cycles (C) were highly significant (P<0.01) for most of these traits. The N x E interactions were significant (P<0.05) for days to mid-silk, lodging and ear acceptability while the C x E interactions were significant for grain yield, mid-silk, and plant height. These significant interactions were due to differences in magnitude rather than the rank order of values of the traits in the different environments. No significant C x N interaction effects were detected for any trait, indicating the magnitude of differences among cycles did not differ between N treatments. TABLE 1. Combined analyses of variance for agronomic traits of cycles 0, 2, 4 and 6 of La Posta maize population measured at 6 locations in Ghana during 1986
* ** Significant at 5% and 1% levels of probability; respectively Since the C x N interaction effects were not significant for all traits, the nature of the N and cycle responses can be entirely explained by their main effects (Table 2). Mean grain yield at zero applied N averaged over replications, environ-ments, and cycles was approximately 37% and 43% lower than grain yield at 80 and 160 kg N ha-1, respectively. Grain yield at 160 kg N ha-1 was approximately 10% higher than yield at 80 kg N ha-1, but the difference was not significant. Grain yields did not differ significantly among cycles 0, 2 and 4, averaging 4.2 Mg ha-1. Cycle 6 significantly out-yielded the other cycles by 9.5%. The significant N x location interactions for grain yield suggests that the yield response to N differed from one location to another. The high-yielding environments were Damongo, Kwadaso and Kpeve and the low-yielding were Ejura, Pokuase and Nyankpala (Fig. 1). Substantial yield increases were observed from 0 to 80 kg N ha-1 in all environments (Fig. 1). These yield increases ranged from 31% in the coastal savanna to 107% in the Guinea savanna. Yield increases of 11%, 15%, 21% and 40% were observed from 80 to 160 kg N ha-1 at Nyankpala, Kwadaso, Pokuase and Ejura, respectively. No further yield increase was detected at Damongo and Kpeve beyond the 80 kg N ha-1 rate. The lowest grain yield (1.5 Mg ha-1) was observed at zero N in the Guinea savanna and the highest (4.4 Mg ha-1) was observed in the forest zone, reflecting the well-known soil fertility differential between the two zones. The relatively high yields observed under unfertilised conditions at some of the locations were attributed to mineralisation of organic residues, and atmospheric N fixation by previous leguminous crops. Figure 1: Grain yield response of La Posta maize population to N fertilizer application at six locations in Ghana, 1986. TABLE 2. Mean response of cycles 0, 2, 4 and 6 of selection in La Posta maize population to 3 levels of N fetlilisation evaluated at 6 sites in Ghana during 1986
+ = no lodging or good ear, 5 = all plants lodged or poor ear Individual cycle response of grain yield to N fertilisation is presented in Table 2. Grain yield of all cycles increased as fertilizer N supply was increased. In all cycles, the response of yield to applied N was greater for the first than the second increment of 80 kg N ha-1. For example, the yield difference between the unfertilised treatment and 80 kg N ha-1 was 1.9 Mg ha-1 compared with 0.6 Mg ha-1 between 80 and 160 kg N ha-1 for the sixth cycle of selection. The yield response was similar for all cycles (Table 2). Nitrogen treatment means for the other agronomic traits showed the response of these traits to N fertilisation was similar at 80 and 160 kg N ha-1 (Table 2). There was a significant delay in silk emergence, a significant reduction in plant height and lodging, but grain moisture and number of ears per plant did not change when N fertilizer was not applied. Ears from plants grown without N fertilisation were rated less acceptable than ears from plants which received fertilizer N. Though silk was delayed by two days when fertilizer N was not applied, no significant differences were observed among N treatments for grain moisture content at harvest. This suggests that grain filling was curtailed at low N, hastening the dry-down of the grain. Consequently, grain moisture was similar for all N treatments. Least square estimates of linear response per cycle of full-sib family selection under the three N regimes and averaged over N treatments for four agronomic traits are presented in Table 3. Since no significant C x N interaction was detected for any of the traits, these estimates of linear changes should be interpreted with caution. Linear gains per selection cycle were highly significant (P<0.01) for grain yield, mid-silk, plant height, and lodging at the 80 and 160 kg N ha-1 rates. Gains were highly significant also for mid-silk and plant height under the unfertilised condition and when estimated over the three N levels. Linear increase in grain yield per cycle was 1.6% at zero-N, 2.1% at 80 kg N ha-1, 1.8% at 160 kg N ha-1, and 1.6% across N levels. The total yield gains from cycles 0 to 6 were 9% under zero-N, 11% under 80 kg N ha-1, 13% under 160 kg N ha-1 and 9% across the three N levels. These results indicate that selection under optimum N-levels during the six cycles resulted in similar yield gains under the three N levels and across N levels. TABLE 3. Progress per cycle of full-sib selection for 4 traits in La Posta maize population evaluated under 3N rates at 6 sites in Ghana during 1986
** Significant at 1% levels of probability Yield gains observed in the present study ranged from 1.6-2.1% per cycle compared with the 2.5% reported in the same population evaluated under optimum N rates (100-200) kg N ha-1 at six sites in South and Central America (Pandey et al., 1986). These rates of linear increase were low compared with the 5.4% average reported in the literature for grain yield in maize (Hallauer and Miranda, 1981), which may be attributed to the rather low selection intensity (approximately 40%) used in the improvement of the La Posta maize population. Comparable progress per cycle of selection was made in reducing plant height and mid-silk at 80 and 160 kg N ha-1, but these gains were less than those reported by Pandey et al. (1986). The greatest gain was made in reducing lodging at 80 kg N ha-1 (3.5%) and at 160 kg N ha-1 (2.8%). No significant response from selection was observed for grain moisture, ear acceptability, and number of ears per plant at both N levels. Pandey et al. (1986) did not report on grain moisture but they also did not observe any significant progress from selection for ear acceptability and number of ears per plant. The data presented seem to suggest that selection under intermediate to high N levels resulted in similar gains under low as well as under high N. These results are in contrast with previous reports on improvement of maize for performance under low N which generally concluded that gains under low N can best be achieved through deliberate selection in low-N environments (Banziger et al., 1997; Banziger and Lafitte, 1997; Lafitte and Edmeades, 1994a, 1994b, 1995; Lafitte et al., 1997). For example, Lafitte and Edmeades (1994a) reported that the largest grain yields in low-N environments were obtained from direct selection for yield under N stress. The relatively high initial soil fertility observed at three out of the six experimental fields in the present study probably resulted in over-estimation of gains under low N. Results from the six cycles studied showed that (i) grain yield increased tremendously over the yield at zero-N following application of 80 or 160 kg N ha-1, (ii) the yield increment from 80 to 160 kg N ha-1 was not significant, (iii) Cycle 6 significantly out-yielded the other cycles across N levels, (iv) full-sib selection was effective in improving grain yield, mid-silk, plant height, and lodging in the population when gains were estimated under moderate to high N levels, and (v) recurrent selection for improved agronomic performance did not significantly alter the N response of the population. ACKNOWLEDGEMENTS We are grateful to the CIMMYT Maize Programme for providing the genetic materials evaluated in the study and to the Ghana-CIDA Grains Development Project and the Ghanaian-German (GTZ) Project at Nyankpala for providing funds and facilities for this study. We gratefully thank Messrs. B.D. Dzah, C.P.K. Dugbenu, E.M. Lampoh, R.K. Kankam, B.D. Dodoo, S.K. Bonsi, M. Awalime, F. Dzoagbe, A.D. Mohammed and I.S. Mohammed, all of the Crops Research Institute of Ghana, for their help in the management of the trials at the various sites. REFERENCES Ahn, P. 1970. West African Soils. Oxford University Press, Ely House, London. 332 pp. Allan, A.Y. and Darrah, L.L. 1978. Effects of three cycles of reciprocal recurrent selection on the N and plant population responses of two maize hybrids in Kenya. Crop Science 18:112- 114. Balko, L.G. and Russell, W.A. 1980. Effects of rates of nitrogen fertilizer on maize inbred lines and hybrid progeny. I. Prediction of yield responses. Maydica 25:65-79. Banziger, M., Betran, F.J. and Lafitte, H.R. 1997. Efficiency of high-nitrogen selection environments for improving maize for low-nitrogen target environments. Crop Science 37:1103-1109. Banziger, M. and Lafitte, H.R. 1997. Efficiency of secondary traits for improving maize for low-nitrogen target environments. Crop Science 37:1110-1117. Carlone, M.R. and Russell, W.A. 1987. Response to plant densities and nitrogen levels for four maize cultivars from different eras of breeding. Crop Science 27:465-470. Castleberry, R.M., Crum, C.W. and Krull, C.F. 1984. Genetic yield improvement of U.S. maize cultivars under varying fertility and climatic environments. Crop Science 24:33-36. Chevalier, P. and Schrader, L.E. 1977. Genotypic differences in nitrate absorption and partitioning on N among plant parts in maize. Crop Science 17:897-901. CIMMYT. 1987. CIMMYT Report on Maize Improvement. CIMMYT, El Batan, Mexico. Dennis, E.A. 1983. Fertilizers for maize on Ghanaian Soils. In: Summarized proceedings 1981-1984, Ghana National Maize Workshop, pp. 84-87. Ghana Grains Development Project, Crops Research Institute, Kumasi, Ghana. Djokoto, R.K. and Stephens, D. 1961. Thirty long-term fertilizer experiments under continuous cropping in Ghana. I. Crop yields and response to fertilizers and manures. Empire Journal of Experimental Agriculture 29:182-195. Hallauer, A.R. and Miranda, J.B. 1981. Quantitative Genetics in Maize Breeding. Iowa State Univ. Press, Ames, Iowa. 468pp. Harvey, P.H. 1939. Hereditary variation in plant nutrition. Genetics 24:437-461. Kamprath, E.J., Moll, R.H. and Rodriguez, N. 1982. Effects of nitrogen fertilization and recurrent selection on performance of hybrid population of Zea mays L. Agronomy Journal 74:955-958. Krantz, B.A. and Chandler, W.V. 1954. Fertilize corn for higher yields. North Carolina Agricultural Experiment Station Bulletin 366. Lafitte, H.R. and Edmeades, G.O. 1994a. Improvement for tolerance to low soil nitrogen in tropical maize. I. Selection criteria. Field Crops Research 39:1-14. Lafitte, H.R. and Edmeades, G.O. 1994b. Improvement for tolerance to low soil nitrogen in tropical maize. II. Grain yield, biomass production, and N accumulation. Field Crops Research 39:15-25. Lafitte, H.R. and Edmeades, G.O. 1995. Stress tolerance in tropical maize is linked to constitutive changes in ear growth characteristics. Crop Science 35:820-826. Lafitte, H.R., Edmeades, G.O. and Taba, S. 1997. Adaptive strategies identified among tropical maize landraces for nitrogen-limited environments. Field Crops Research 49:187-204. Lugg, D.G. and Sinclair, T.R. 1981. Seasonal changes in photosynthesis of field-grown soybean leaflets. 2. Relation to nitrogen content. Photosynthetica 15:138-144. Moll, R.H., Kamprath, E.J. and Jackson, W.A. 1987. Development of nitrogen efficient prolific hybrids of maize. Crop Science 27:181-186. Muchow, R.C. 1988a. Effect of nitrogen supply on the comparative productivity of maize and sorghum in a semi-arid tropical environment. I. Leaf growth and leaf nitrogen. Field Crops Research 18:1-16. Muchow, R.C. 1988b. Effect of nitrogen supply on the comparative productivity of maize and sorghum in a semi-arid tropical environment. III. Grain yield and nitrogen accumulation. Field Crops Research 18:31-43. Murata, Y. 1969. Physiological responses to nitrogen in plants. In: Physiological Aspects of Crop Yield. Eastin, J.D, Haskins, F.A., Sullivan, C.Y. and van Bavel, C.H.M. (Eds.), pp. 235-263. American Society of Agronomy, Madison, Wisconsin. Muruli, B.L. and Paulsen, G.M. 1981. Improvement of nitrogen use efficiency and its relationship to other traits in maize. Maydica 26:63-73. Novoa, R. and Loomis, R.S. 1981. Nitrogen and plant production. Plant Soil 58:177-204. Pandey, S., Diallo, A.O., Islam, T.M.T. and Deutsch, J. 1986. Progress from selection in eight tropical maize populations using international testing. Crop Science 26:879-884. Pollmer, W.G., Eberhard, D., Klein, D. and Dhillon, B.S. 1979. Genetic control of nitrogen uptake and translocation in maize. Crop Science 19:83-86. Sanchez, P.A. 1976. Properties and Management of Soils in the Tropics. Wiley-Interscience Publication, John Wiley & Sons, New York, USA. 618pp. Steel, T.G. and Torrie, J.H. 1980. Principles and Procedures of Statistics: A Biometric Approach. McGraw-Hill Book Company Inc., New York, USA. 568pp. Viets, F.G. 1965. The plant's need for the use of nitrogen. In: Bartholomew, W.V. and Clark, F.E. (eds.), Agronomy 10:503-549. Copyright 1998, African Crop Science Society The following images related to this document are available:Line drawing images[cs98026a.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}