 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 6. No. 4, pp. 329-344,
1998 Performance Of Early Maturing Katumani And Kito Maize Composites Under Drought at the Seedling And Flowering Stages S.N. MUGO, M.E. SMITH1, M. BäNZIGER2, T.L. SETTER3, G.O. EDMEADES and A. ELINGS International Maize and Wheat Improvement Centre (CIMMYT),
Lisboa 27, Apdo Postal 6-641, 06600 México, D.F.,
México (Received 23 April, 1998; accepted 29 September, 1998) Code Number:CS98035
ABSTRACT Two widely-used open pollinated maize (Zea mays L.) cultivars from East Africa, Katumani and Kito composites, developed and improved for early maturity, grain yield and drought escape were evaluated for drought tolerance at seedling and flowering stages, and compared with other drought tolerant maize populations. Under seedling-stage, the early maturing cultivars were the most drought tolerant with large biomass accumulation in both stress and non-stress environments. However, root weight and the root: shoot ratio were relatively small. Under drought at flowering, these cultivars flowered early and reached anthesis in 66 days (d) for Katumani and 73 d for Kito, compared to an average of 100 d for the trial. Grain yields were (359 kg ha-1 for Katumani and 679 kg ha-1 for Kito) than the trial average of 1002 kg ha-1 under severe stress. Under well-watered conditions the yields were 3,295 kg ha-1 for Katumani, 4,651 kg ha-1 for Kito compared to the trial average of 5,737 kg ha-1. Low grain yield was associated with a large anthesis-silking interval (ASI) of 28 d in Katumani compared to an average of 18 d for the trial; and strong leaf rolling and high leaf abscisic acid concentration. Katumani and Kito were less stable for yield across water availability environments. Key Words: Abscisic acid, anthesis-silking interval, drought tolerance, root:shoot ratio, yield stability RÉSUMÉ Deux cultivars de maïs ouverts et fécondés largement employés dans les régions centrales et australes de l'Afrique, composés Katumani et Kito, ont été développés et améliorés pour la maturité précoce; le grain de production et l'évasion à la sécheresse ont été évalués pour la tolérance à la sécheresse aux niveaux de semis et de la floraison, et comparés à d'autres populations de maïs tolérantes à la sécheresse. Pendant les semis, les cultures à maturité précoce ont été les plus tolérantes avec une large accumulation de la biomasse dans les milieux stressés et non stressés. Cependant, le poids de racines et le rapport racine/tige était relativement petit. Pendant la sécheresse, à la floraison, les cultures avaient fleuri bien avant et ont atteint l'anthèse à 66 jours pour le Katumani et 73 jours pour le Kito, comparés à une moyenne de 100 jours par essai. Les grains de production ont été de 359Kg ha-1 pour le Katumani et de 679Kg ha-1 pour le Kito tandis que la moyenne d'essai était de 1002Kg ha-1 sous une sévère contrainte. Dans des conditions de bonne irrigation, les rendements étaient de 3.295Kg ha-1 pour le Katumani, 4.651Kg ha-1 pour le Kito, par rapport à la moyenne d'essai de 5.737Kg ha-1. La faible production du grain était associée à une large intervalle d'ASI de 28 jours pour le Katumani par rapport à une moyenne de 18 jours pour l'essai; et a` de grandes et longues feuilles enroulées avec une concentration d'acide abiscique. Le Katumani et le Kito étaient moins stables pour la production dans les milieux humides. Mots Clés: acide abiscique, intervalle anthèse, siking, tolérance à la sécheresse, racine: rapport de la pousse, stabilité rendement INTRODUCTION Drought stress at seedling and flowering stages of maize (Zea mays L.) is estimated to cause annual yield losses of about 17% in the tropics (Edmeades et al., 1994). Complete crop failure, reduced establishment, or reduced yield may result if drought occurs at the seedling, flowering or grain filling stages. Eastern Africa has recently experienced periodic food shortages attributable, in part, to intermittent but severe droughts (FAO, 1997). High population pressure in the East African region, which is expected to double in 20 years, is forcing more intensive cultivation of existing farms and movement into previously uncropped areas. Poverty and famine, especially that induced by drought, often prevent farmers from adopting conventional management practices because of capital constraints and risk. Given that favorable land is already almost fully utilised, future production increases will come only from boosting yields or from expanding maize production into less favorable areas (KARI, 1994). Stress tolerant maize varieties offer a means of stabilising yields at no additional cost to the farmer (Edmeades et al., 1997). Maize is the principal cereal crop in East Africa. It ranks first in total production and yields per hectare in Ethiopia (Mohammed and Mulatu, 1993) and is the most important food crop grown and consumed in Tanzania (Mduruma and Ngowi, 1997) and Kenya (KARI, 1994). In Somalia, maize ranks second after sorghum in acreage. Kenya, Tanzania and Ethiopia each plant more than a million hectares of maize annually, and Burundi, Somalia, and Uganda each sow more than 100,000 ha (CIMMYT, 1994). Shortages in maize production in the region occur periodically, necessitating costly alleviation through food aid. One way to improve the livelihood of small holders is to reduce the fluctuation in maize production over the region in the face of varying annual rainfall. Decreasing the susceptibility of maize to drought, while maintaining or increasing yield in good rainfall years, would increase and stabilise rural incomes, reduce chronic food shortages that these areas experience prior to harvest, and lessen the risk of famine. Quantification of occurrence and timing of drought for six maize agro-ecologies in Kenya revealed the presence of drought during the seedling stage (the first 10-20 d after rainfall onset) in all zones, and during flowering and grain filling stages (Njoroge et al., 1997). These authors expressed need for drought tolerant maize germplasm in four of the six agro-climatic zones, through exposing breeding materials and segregating progenies to drought stress during periods of crop development. In Tanzania, where maize losses up to 50% due to drought stress are common, early maturing maize populations have been developed, but are very sensitive to early season stress (Mduruma and Ngowi, 1997). Maize has become important in Ethiopia, where moisture stress is a constraint on about 40% of the cropped area (Mohammed and Mulatu, 1993). Early maturing maize cultivars perform well in the Rift Valley region of Central Ethiopia (Ransom et al., 1997; Yitbarek, 1997). Late maturing drought tolerant cultivars are necessary in that region, especially if they have the ability to bridge a drought period between the two major rainy seasons (Ransom et al., 1997). Two major categories of drought resistance can be distinguished: drought escape and drought tolerance (Blum, 1988). Under drought escape, the plant completes critical physiological processes before drought sets in, while drought tolerance implies the presence of physiological mechanisms that allow the plant a reasonable level of production despite the presence of drought (Blum, 1988). Short season, early maturing cultivars frequently escape drought that occurs at either end of a rainy season, but normally have limited yield potential (Blum, 1988). Escape through early maturity is rarely sufficient in East Africa since the occurrence of drought is erratic, often varying in intensity and timing; it occurs at any stage of growth. One approach to improving crop performance in water-limited environments is to select for genotypes that have improved yield in these environments (Mullet and Whitsitt, 1996). Another approach involves the identification and selection of secondary traits that contribute to drought avoidance, drought tolerance, or water use efficiency (Ludlow and Muchow, 1990). Water status traits like leaf abscisic acid concentration, osmotic adjustment, water uptake, and leaf waxiness (among others) could potentially be useful in selection for seedling stage drought tolerance. The phytohormone, ABA, is ubiquitous in all flowering plants. When plants wilt, ABA levels typically rise as a result of an increase in the rate of synthesis (Taylor, 1991). Increasing ABA concentration leads to many changes in crop physiology, growth, and development. Responses to increased ABA levels include reduced leaf area development (Hall and McWha, 1981), enhanced root:shoot ratio (Saab et al., 1990; Hall and McWha, 1981), reduced stomatal conductance and plant transpiration rates (Koch and Bergman, 1979). All are usually in a consistent direction that would favour plant adaptation to drought (Jones, 1979). Stomatal control and leaf rolling allow short-term and reversible regulation, leading to a reduced water flux and consequently to a decreased difference between root and leaf water potentials. This short-term regulation is usually not sufficient, so plants change their architecture by reducing leaf growth, increasing root:shoot ratio, and reducing leaf area by early senescence (Tardieu, 1996). Constitutive traits, such as seed size, may also influence seedling establishment under drought stress (Mian and Nafziger, 1994). We still lack an understanding of how seed and plant water status interact with the processes of germination, emergence and establishment. When drought stress occurs just before or during the flowering period in maize, a delay in silk emergence is observed resulting in an increase in the length of the anthesis-silking interval (Bolaños and Edmeades, 1993). This asynchrony between male and female flowering dates is strongly associated with grain yield decrease under drought conditions. Selection for reduced anthesis- silking interval (ASI) in tropical open-pollinated varieties has been shown to correlate well with improved number of ears per plant and grain yield under drought stress (Bolaños and Edmeades, 1993). Delayed leaf senescence under drought increases the period over which photosynthesis can take place. Delayed leaf senescence may occur as a result of direct genetic controls or it may indicate access to a larger amount of water (deeper rooting), efficient regulation of water loss by the genotypes, or osmoprotection via leaf osmotic adjustment. Populations with tolerance to seedling or flowering drought stress have been developed at the International Maize and Wheat Improvement Center (CIMMYT) using carefully managed drought stress in the field (Edmeades et al., 1997). Seedling survival, biomass production, unrolled leaves under drought, and recovery after irrigation are the morphological traits used as main selection criteria for seedling drought tolerance at CIMMYT (Bänziger et al., 1997). Selection for tolerance at flowering was for an index of traits that sought to increase grain yield, reduce ASI, increase ears per plant, delay leaf senescence, reduce tassel size, and increase leaf angle while leaving male flowering date unaltered. Traits related to yield and flowering received the greatest weighting. Efforts to breed specifically for drought tolerance have been limited in the East African region. Most programmes have either concentrated on developing cultivars that can escape drought through earliness (Mugo and Njoroge, 1997), focus on more favored environments (Mduruma and Ngowi, 1997), or test Afinished@ varieties and hybrids in dry zones to evaluate their response (Mugo and Njoroge, 1997; Ransom et al., 1997; Yitbarek, 1997). However, transfer of improved germplasm developed in México or Southern Africa is limited by adaptation to other biotic and abiotic stresses unique to the East African environment. Sources of tolerance are best sought within materials adapted to the region, before introgression of exotic sources. The objectives of this study were to evaluate the potential of Katumani and Kito composites as sources of germplasm for tolerance to drought and to identify avenues for their further improvement. MATERIALS AND METHODS Genetic materials. Twenty maize cultivars were evaluated at seedling, flowering and grain filling stages. These included three cycles resulting from S1 recurrent selection for flowering and grain filling drought tolerance from four population groups, namely; Tuxpeño Sequía, La Posta Sequía, Pool 26 Sequía, and Pool 18 Sequía. Eight additional populations were included as in Table 1. Only contrasts of interest to East Africa, which include the latest cycle of each of the four population groups presented here. Katumani and Kito are two early maturing East African cultivars being grown extensively by farmers; and TS6, La Posta Sequía, Pool 26 Sequía and Pool 18 Sequía, DTP2 W and Pool Latente are eight populations from CIMMYT in México. These cultivars are source germplasm with tolerance to mid-season drought, adapted to the tropics. The drought tolerant population 1 (DTP1) selections: seedling best all (DTP1 SIBA), and seedling worst all (DTP1 SIWA) are divergent selections for best and worst survival and seedling performance, under drought stress imposed during crop establishment (Bänziger et al., 1997). TABLE 1. Descriptions and pedigrees of ten tropical maize cultivars evaluated at seedling, flowering and grain filling stages under a range of soil moisture status in a greenhouse at Ithaca, NY, USA, and in the field at Tlaltizapan, Mexico
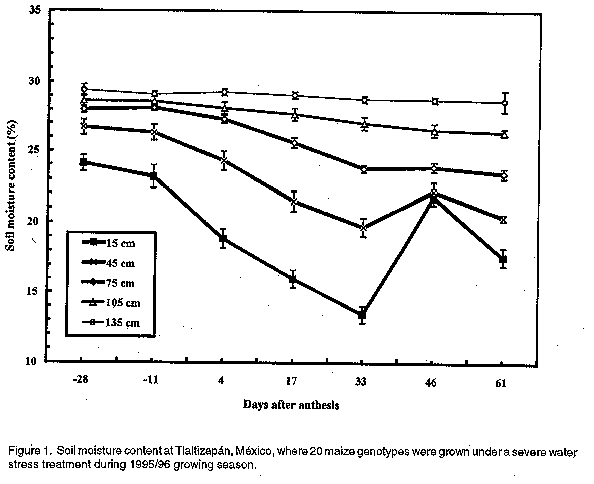
Seedling stage greenhouse evaluation. Seedlings were grown in two seasons during May-July and November-December 1996 in a greenhouse, Cornell University, Ithaca, NY, USA, under 24º C day and 20º C night temperatures, 75% relative humidity, and a 14 hour day 100-watt metal halide lamps to boost winter radiation levels supplemented solar radiation. Plants were exposed to two water availability treatments, viz. severe stress and well watered. Water was applied to field capacity in the pots but ceased 10 d after emergence in the severely stressed treatment. Cultivars in both treatments were separately sown in a randomised complete block design with three replications. Five plants per pot were grown in 23 cm diameter plastic pots, each plot comprising two pots. Cornell mix, a mixture of peat moss and vermiculite, a Peters unimix plus II 10-10-5 formula (Booley and Sheldrake, 1977) was used with 15 grammes per pot of supplemental iron (III) sulphate powder. Fertilizer was added at the rate of 0.3 g each of elemental N and K, and 0.1g P per pot 10 d after sowing. The well-watered treatment received additional fertilizer at the same rate 14 d after sowing. Seedling vigour, a visual evaluation of biomass production per pot, was scored once at 25 d after sowing during the first season, and three times at 18, 25, and 47 d after sowing during the second season. Scoring was on a 1-5 scale (1= least vigorous, 5= very vigorous). Leaf rolling was scored 26 and 30d after sowing in the first season, and 18 and 47d after sowing during the second season on a 1-5 scale (1=least rolled, 5=very rolled). The number of senescent leaves for each plant was recorded at 38 and 44d after sowing during the second season only. These scores or counts were averaged over time within each season. Leaf disks for ABA assay were sampled 25 and 36 d after sowing (designated Aearly-leaf ABA concentration@ and Alate-leaf ABA concentration@, respectively). One leaf disc sample was taken from each of the 10 plants per plot, halfway along the length of the most recently fully expanded leaf but avoiding the midrib. ABA was extracted with 80% methanol with 1% (volume basis) glacial acetic acid. ABA was assayed using a modified ELISA method as described by Ober et al. (1991). The number of fully expanded leaves was counted at harvest time (the first season at 35 and the second season at 47d). Plant height was measured at harvest as the distance from pot soil level to the tip of the most recently fully expanded leaf for all plants in the pot. Shoot and root weights were measured after harvesting and drying the materials in a hot air oven at 60 0 C for 10 d. A sample of seeds from the same seed lots used in these studies was weighed and 100-seed weight calculated. Flowering and grain filling stage field evaluation. Twenty cultivars were grown during the rain-free winter seasons of November-April 1995/96 and 1996/97 at CIMMYT's experiment station at Tlaltizapán, Morelos, México (190N, 940 m above sea level). Data from only the ten cultivar contrasts described above were used for this report. The mean maximum and minimum temperatures during the winter growing season at this location are 32 0C and 14 0C, respectively (Chapman, 1995). A 4x5 alpha (0,1)-lattice design with 3 replications was used. Plots consisted of 4 rows of 5 meter length, with 75 cm spacing between and 25 cm within rows. Two seeds per hill were sown and later thinned to one to give the desired density of 53,000 plants ha-1. Cultivars were sown at different times in an attempt to have all cultivars reach anthesis at the same time. Due to the limitations imposed by irrigation, two separate but adjacent fields were planted each for the severe stress and well-watered treatments. Irrigation was by gravity flow in open furrows. In the severe stress treatment, irrigation decreased gradually to ensure that drought stress peaked at flowering. Adequate watering every 10-12 d was maintained in the well-watered treatment. Some irrigation was applied to the severe stress treatment 120 d after sowing of the late flowering cultivars, to ensure grain filling. Soil moisture content was measured with a neutron probe (Model 503DR, CPN Company, 2830 How Rd, Martinez, CA 94553, USA) every two weeks starting from when irrigation was initially withdrawn in the severe stress plots. Four tubes were installed per replication and soil moisture content measured at 15, 45, 75, 105 and 135 cm depths. During tube installation, samples from the removed soil cores were oven-dried and used to calibrate probe readings for moisture content. The soil moisture contents estimated by the probe were averaged for the individual depths and periods of measurements. Leaf rolling was scored as in the seedling drought evaluation trial when symptoms were apparent. Three scores were taken prior to flowering, and an average computed. Leaf senescence was estimated from three independent scores taken in the last half of grain filling on a 0-10 scale (0 = 0% dry leaves, 10 = 100% dry leaves). Leaf ABA concentration was determined from assays as previously described. Samples were taken just prior to anthesis. One leaf disc from each of 12 plants in the nets plot was taken from the middle of the blade of the flag leaf, or from the most recently fully expanded leaf. Plant and ear heights were measured from ground level to the base of the flag leaf and the primary ear, respectively. Measurements were taken from six well bordered plants and an average calculated. Root lodging was determined as the number of plants whose stems were inclined beyond 300 from the vertical in the plot. Days to 50% anthesis and silking were estimated from regression of the linear phase of daily accumulated totals of flowering plants on time from sowing, estimated on a per plot basis. Days to 50% anthesis or silking were estimated by equating Y value to half the number of plants in the plot. The anthesis-silking interval was calculated as the difference in time between 50% silking and 50% anthesis. The number of plants and ears harvested in the two middle rows per plot after the end plants had been removed as borders were counted (net harvested area was 5.63 m2) and ears per plant calculated. Shelled grain weight was expressed on oven-dry basis. Statistical analysis. Data obtained were subjected to analyses of variance. Genotypes and water treatments were considered fixed effects, while seasons and years were considered random effects. Homogeneity of variances from the two years' data was confirmed and a combined analysis over water treatments and seasons or years' performed, and F-tests for statistical significance conducted. Standard errors and Fisher=s least significant difference among means for each trait were computed. Correlation coefficients among traits were also calculated. The Eberhart and Russell (1966) model for stability analysis was used to estimate regression coefficients (bi) of yield of individual cultivars in the four environments where each was grown. Regression coefficients (bi) were tested for significant differences from unity using t-tests, while the significance of the deviations from regression (S2di) were tested by the F-test based on pooled error estimates. RESULTS Seedling stage. Combined analysis of variance indicated significant differences due to water treatment, season, and season x water treatment interactions, for all traits (Table 2). Cultivars were significantly different only in seedling vigour and total dry weight. There were no significant cultivar x water treatment or cultivar x season interactions, but there were significant cultivar x water x season interactions for all traits measured. TABLE 2. Sum of squares from ANOVA for traits measured in 10 maize cultivars grown at seedling stage under severe water stress and in a well-watered treatment imposed in a greenhouse over two seasons1
TABLE 2 Continued:-
1 The two seasons of seedling evaluation were two sets grown at
different times in summer and fall, 1996 Leaf rolling and senescence, root weight, root:shoot ratio, and leaf ABA concentrations were significantly greater (P<0.01) in the severe stress treatment compared to the well-watered treatments (Table 3). The number of fully expanded leaves, seedling height, shoot weight, and total dry weight were significantly reduced by the severe stress compared to the well-watered treatment (Table 3). Drought tolerance was assumed to be related to genotypic ability to accumulate high total dry weight under drought stress. Total dry weight for Katumani and Kito cultivars were approximately the average of the cultivars evaluated under the severe stress treatment. In the well-watered treatment, Katumani had the greatest total dry weight. Both composites had high seedling vigour scores under the severe stress treatment, matched only by the vigour of Pool Latente (Table 3). Katumani=s seedling vigour score was also high in the well-watered treatment, as were those of Kito, La Posta Sequía and Pool Latente. Katumani seedlings were tall, second only to those of Pool 18 Sequía, while Kito seedlings were much shorter. Katumani seedlings were significantly taller than all others under the well-watered treatment. TABLE 3. Means of traits measured on 10 maize cultivars at seedling stage when grown under severe water stress or in a well-watered regime in a greenhouse, over two seasons1
TABLE 3 Continued:
Leaf rolling scores under severe stress for Katumani were higher than for Kito (Table 3). Stress even at the seedling stage induced leaf senescence, and Kito had a higher leaf senescence score than Katumani. This ranking was similar to the well-watered treatment. There were no significant differences in number of fully expanded leaves among cultivars in either the severe stress or well-watered treatments. Katumani had the highest leaf ABA concentration under both the severe stress and well-watered treatments. Kito had above average leaf ABA concentration under severe stress but a very low concentration under the well-watered treatment (Table 3). Under severe stress, Katumani and Kito had average root dry weights while Pool Latente had the highest root weight (Table 3). In the well-watered treatment, Katumani had the greatest root dry weight. Shoot dry weights for Katumani and Kito were under severe stress, but Katumani had the highest shoot dry weight under the well-watered treatment. Katumani, therefore had a markedly lower root:shoot ratio under severe stress compared with other cultivars, though under the well-watered regime, Katumani=s root:shoot ratio was still relatively high. While root:shoot ratio was higher under severe stress than under well-watered conditions for all cultivars, Katumani, TS6, Pool 18 Sequía, and DTP1 SIWA developed relatively more shoot at the expense of roots (root:shoot ratio was less than 0.50), while Kito was relatively lower for this trait. Under severe stress, there were no significant differences between DTP1 SIWA and DTP1 SIBA in seedling vigour, shoot dry weight, seedling heights, and leaf ABA concentration. DTP1 SIWA was significantly greater than DTP1 SIBA in leaf rolling scores, but significantly less in leaf senescence, root weight, and root:shoot ratio. Under the well-watered regime, DTP1 SIWA had significantly greater seedling vigour, leaf senescence, root weight, shoot weight and total dry weight than DTP1 SIBA. Flowering and grain filling stage. Soil moisture content decreased significantly between the surface and 135 cm depth in the severe stress treatment during 1995/96 season (Fig. 1). Figure 1. Soil moisture content at Tlaltizapan, Mexico, where 20 maize genotypes were grown under a severe water stress treatment during 1995/96 growing season. Anthesis and silking occured on average at 82 d and 100 d after sowing, respectively. The cultivars, therefore, experienced drought stress at flowering. Cultivars acquired most of their water by depleting it from depths less than 75 cm; there was available water deeper in the profile even at harvest time. Drought stress may have limited deep rooting, and deep-rooted cultivars could have benefited from water held below 75-cm depth. Analyses of variance combined across years and water treatments indicated significant (P<0.05) differences between water treatments for all measured traits except anthesis and leaf ABA concentration (Table 4). Cultivars showed significant (P<0.05) differences except for root lodging and leaf senescence scores. There were significant differences between years, except for ears per plant, days to silking and ASI. There were no significant cultivar x water interactions; only leaf ABA concentration showed a cultivar x year interaction. However, there were significant cultivar x year x water interactions for all traits with the exception of root lodging, leaf senescence, ABA concentration and ASI. As in the seedling trial, therefore, conclusions are drawn from data analysed by each water treatment across the two years. TABLE 4. Sum of squares for traits when a severe moisture stress or no stress was imposed on 10 maize cultivars at the flowering and grain filling stages in a field trial conducted over two years at Tlaltizapan, Mexico
TABLE 4 continued:-
1 Leaf senescence scores: (0=0% dry leaves, 10=100% dry
leaves) The presence of significant cultivar x year x water interactions indicated that cultivars responded differently to the specific water treatments in each year. The environments were classified and ordered according to the mean grain yield of all cultivars in that environment. Mean grain yields for these environments were 396 kg ha-1 for SS-96, 1608 kg ha-1 for SS-97, 4355 kg ha-1 for WW-997 and 7119 kg ha-1 for WW-96. Eberhart and Russell (1966) defined stable cultivars as ones with high average grain yields, with regression coefficients less than or equal to unity (bi < 1) indicating high stability over locations, and those with small deviation from regressions (S2di) i.e., those with high coefficient of determination (R2). The coefficients of determination (R2) values were high, indicating that a sizable amount of variability should be attributed to cultivars. Katumani, Kito, Pool 18 Sequía and DTP1 SIBA had regression coefficients below one, which indicated smaller response in grain yields with increasingly favorable environments (Table 5). Katumani and Kito had the lowest average grain yields in all environments, and demonstrated a reduced capacity to respond to good environments. All cultivars evaluated had non-significant deviations from regression (S2di), indicating that the responses of the cultivars to changes of environment fitted a linear regression model. Katumani and Kito had regression coefficients less than unity (P<0.01). These cultivars, therefore, had relatively poor responses to favourable environments. Pool Latente had a regression coefficient significantly (P<0.01) greater than unity, indicating that it was very responsive to favourable environments. DTP1 SIBA, Pool 18 Sequía, TS6 Sequía, and DTP1 SIWA were the most stable cultivars. They had high mean grain yields across environments, regression coefficients close to unity, and small deviations from regression. Pool 26 Sequía, La Posta Sequía and DTP2-W were less stable, indicating good responses to favourable environments but poor responses to unfavourable ones. TABLE 5. Mean grain yields regression coefficients (bi), deviations from regression (S2di), and coefficients of determination (R2 ) of 10 maize cultivars grown under four environments created by subjection to varying levels of water deficits during 1996 - 1997 at Tlaltizapan, Mexico
*, ** Significant at the 5, and 1% levels of probability, respectively Under severe stress, leaf senescence, days to silking, and ASI increased significantly, while plant height, ear height, ears per plant, ear weight and grain yield decreased significantly relative to those of the well-watered treatment (Table 6). Katumani, Kito and Pool 26 Sequía had relatively low grain yields under both the severe stress and the well-watered treatments. Kito did not differ significantly from La Posta Sequía under the severe stress treatment. The low yields in Katumani are associated with a small number of ears per plant and a reduced ear weight in both environments. Kito had a larger number of ears per plant, but smaller ear weight under the severe stress treatment. Barrenness in Katumani therefore, increased dramatically under the severe stress. Since weights per kernel for Katumani and Kito were high (Table 3), their low grain yields reflect fewer kernels produced under both the severe stress and well-watered treatments. The divergent selections for seedling survival under drought stress, DTP1 SIWA and DTP1 SIBA were not significantly different for grain yield under both severe stress and well-watered treatments. TABLE 6. Means of traits measured from 10 maize cultivars subjected at flowering and grain filling stages to a severe water stress or a well-watered regime at Tlaltizapan, Mexico, over two years
TABLE 6 continued:-
1 Leaf rolling scores: (1=L east rolled, 5= Very rolled). Score
takes into account extent of rolling of individual leaves and
percentage of leaves rolled Katumani and Kito cultivars had the shortest interval from sowing to anthesis under both water treatments. However, under severe stress their time to silking were considerably increased, thus significantly increasing ASI. Pool 26 Sequía had the greatest ASI followed by Katumani, while Kito had below average ASI under the severe stress treatment. Leaf rolling scores under severe stress were the highest in Katumani, while Kito came second in a group that included Pool 26 Sequía, Pool 18 Sequía, and Pool Latente. Katumani also had a high leaf senescence score, second only to Pool 26 Sequía under severe stress, but had a very low leaf senescence score under the well-watered treatment. Kito had average leaf senescence score under severe stress, but a high leaf senescence score when well watered. Katumani had a high leaf ABA concentration under the severe stress comparable to those of Pool 18 Sequía and DTP2 W. Leaf ABA concentration of Kito was average for the severe stress treatment and the well-watered treatment. Katumani and Kito showed the greatest incidence of root lodging under severe stress but were average for this trait under well-watered conditions. The increased root lodging under the well-watered treatment in all cultivars was due to a severe storm that occurred just prior to harvesting in the 1997 season. Under severe stress, Katumani and Kito composites had similar plant heights to those of the other cultivars, but both had significantly lower (P<0.05) ear heights. Katumani, Kito, and Pool 18 Sequía were the shortest in plant and ear heights under the well-watered treatment. Early maturity and increased vigour were advantages for these cultivars under severe stress where they grew relatively taller, despite being sown later. DISCUSSION Seedling stress tolerance. The high early-season growth rate of Katumani and Kito were evident in both environments, as indicated by seedling vigour, tall seedlings, large shoot weights, and large total dry weight. Vigorous early growth may be related to the large seed size of these cultivars, as reported by Bänziger et al. (1997) in seedlings under field conditions. Katumani and Kito had high leaf rolling scores, low leaf senescence, and high leaf ABA concentration. As total dry weights for these varieties were high, we can assert that these traits did not have negative effects on biomass development. Leaf rolling and high leaf ABA concentration in fact, may have limited water loss via transpiration; while reduced leaf senescence ensured availability of adequate green leaf area for photosynthesis. Katumani and Kito had relatively low root dry weights and, consequently, a low root:shoot ratio under the severe stress treatment. Root:shoot ratios were, however, significantly decreased under the well-watered treatment. This indicated that these early maturing cultivars develop shoot biomass rapidly at the expense of root biomass under severe stress. We hypothesize that increased root weight of Katumani while maintaining its earliness, could improve drought tolerance at the seedling stage. High leaf ABA concentrations were observed in Katumani in both environments. Under severe stress, the vigorous Katumani plant may have depleted water faster from the soil, leading to decreased turgor in the plant, and starting a cascade of events that led to increased leaf ABA concentration (Pierce and Rascke, 1980). However, its large leaf ABA concentration in the well-watered treatment indicates its considerable potential for ABA production. DTP1 SIWA had reduced root dry weight under severe stress; but in the absence of stress, DTP1 SIWA accumulated significantly more total dry weight than DTP1 SIBA. The differences between these two were marginal, though significant, indicating that we stressed the plants more severely than the point at which we could determine seedling survival. If we related drought tolerance at the seedling stage with high total dry weight produced under stress, then Pool 18 Sequía, Kito, Pool Latente, La Posta Sequía, and Katumani are the leading cultivars. Three of these, Katumani, Kito, and Pool 18 Sequía were early maturing. These results indicate that early maturing cultivars were better adapted to seedling drought stress, or they exhibited traits indicative of seedling drought tolerance. Tolerance to drought at the flowering and grain filling stages. The significant differences in the environmental variables: years and year x water effects indicates that we could not create exactly similar drought environments from year to year. The severe stress trial was planted in the same field both years, while the well-watered one was grown in different soil types in the two years. Variations in climatic conditions between years, as well as timing of watering and amounts of water dispensed in an open furrow irrigation system may have been sources of year-to-year differences. Care should, therefore, be taken in creating drought environments in the field conditions for selection, and in interpreting their effects.. Grain yield and number of ears per plant were inherently smaller in Katumani and Kito compared to cultivars that had undergone selection for drought tolerance. Mduruma and Ngowi (1997) reported that low grain yields associated with small ears and small grains were the reasons for limited acceptance of early maturing maize cultivars by farmers in Tanzania. Our results here indicate that a combination of barrenness and floret and kernel abortion is responsible for the low grain yields in Katumani and Kito composites rather than small grain size. It is apparent that the yield potentials of Katumani and Kito are low due to their early maturity, long ASI, coupled with decreased leaf area because of increased leaf rolling, leaf senescence and stomatal closure associated with increased leaf ABA concentration. The large ASI and low grain yield even under well-watered condition attest to the inherently low yield potential and lack of mid- season drought tolerance of Katumani and Kito composites. However, there were no other cultivars in this study that were earlier maturing than these two, and under severe terminal drought this would bring definite advantages. Katumani and Kito showed the lowest mean grain yield and the lowest responses to favourable environments, features that indicate less stability according to Eberhart and Russell (1966). Katumani and Kito would be the cultivars of choice in a region that receives low rainfall in all seasons. However, drought, especially in East Africa is of erratic occurrence and favourable seasons are often interspersed with unfavourable ones (Stewart, 1984; Keating et al., 1992; Njoroge et al., 1997). A cultivar with greater average grain yields and is more responsive to favorable environments would maximise grain yields over seasons in the East African region. We believe that there is room to increase yield potential and tolerance to mid-season stress in Katumani and Kito cultivars even under the short cropping seasons to which they are adapted, by recurrent selection directed at decreasing ASI and leaf rolling. Both should result in a reduction in leaf ABA concentration. As Katumani and Kito are genetically variable open-pollinated composites, we expect genetic variability for these traits to be adequate to support gains from a recurrent selection. However, the inherent low yield potential could be improved by introgression of germplasm from some of the populations evaluated here. According to Grafius (1981) exotic germplasm should only be used as a backup genetic resource for future use, and should be introgressed into adapted cultivars only if such adapted germplasm has inadequate levels of the resistance in question. Katumani and Kito populations exhibited low drought tolerance at the flowering/ grain filling stage, and introgression of germplasm carrying this tolerance would be appropriate. Pool 18 Sequía would be a good source of increased grain yield potential under unfavourable environments without sacrificing early maturity. TS6 could be included as well to maintain the white semi-dent kernel types of these two materials, while DTP2 W would contribute towards high grain yields under favourable environments. Cultivars other than Katumani and Kito, had undergone recurrent selection that involved reducing ASI under drought. Reduced ASI has been associated with altered partitioning of photosynthates that favours growth of the ear under drought, thereby reducing barrenness and increasing harvest index (Bolaños and Edmeades, 1993). DTP1 SIWA was shown to be poor in seedling survival (Bänziger et al., 1997) and would not be suitable, therefore, for introgression. The Pool Latente appears to be a good source of drought tolerance during flowering and grain filling, but may lack requisite disease resistance for stable performance. CONCLUSION Katumani and Kito showed advantages under seedling drought stress, as evidenced by high dry weight accumulation. These early maturing cultivars could have tolerance for seedling drought enhanced by increased root dry weight. Katumani and Kito composites had the lowest grain yield under both stress and non-stress conditions. Improvement for drought tolerance through escape by early maturity, therefore, limited their yield potential under favourable environments. Furthermore, these two cultivars are quite susceptible to drought stress at flowering. Improvement of tolerance at flowering might widen their zone of adaptation, as Katumani and Kito were developed for growing in areas that experience drought stress that occurs with increased frequency at the end of the season, but are now being grown in areas that experience flowering and grain filling drought stress. Katumani and Kito are well adapted to East African environments and diseases (Muhammed and Mulatu, 1993; Mduruma and Ngowi, 1997; Mugo and Njoroge, 1997), and incorporation of flowering drought tolerance would be desirable. Recurrent selection under managed drought and directed towards reducing ASI, leaf senescence and leaf rolling would be expected to increase grain yield and stability. Introgression of drought tolerant germplasm such as that evaluated here, could further enhance flowering and grain filling drought tolerance. ACKNOWLEDGMENTS This work was made possible with facilities provided by Cornell University and CIMMYT, and with financial support from fellowship funding for SNM from the Rockefeller Foundation. Thanks are extended to the Director, Kenya Agricultural Research Institute for granting SNM study leave. Special thanks go to Sr. Juan Carlos Bahena and his team at Tlaltizapán, and the greenhouse staff, Department of Plant Breeding, Cornell University, for their assistance in data collection. This paper is part of a dissertation submitted by SNM in partial fulfillment of the requirements for the Ph.D. degree, Department of Plant Breeding, Cornell University, Ithaca, NY 14853. REFERENCES
Copyright 1998, African Crop Science Society The following images related to this document are available:Line drawing images[cs98035a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}