 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 6. No. 4, pp. 377-383,
1998 Effect Of Soybean, Cowpea And Groundnut Rotations On Root-Knot Nematode Build-Up And Infestation Of Dryland Maize H.F. RIEKERT and G.E. HENSHAW1 ARC-Grain Crops Institute, Private Bag X1251, Potchefstroom,
South Africa (Received 20 October, 1997; accepted 9 September, 1998)
Code Number:CS98040
ABSTRACT The effect of soybean (Glycine max L. Merr), cowpea (Vigna unguiculata L. Walp) and groundnut (Arachis hypogea L.) rotations with maize (Zea mays L.) on root-knot nematode infestation levels was investigated in a crop rotation trial conducted over four seasons. The most prevalent root-knot nematode species were Meloidogyne javanica (Treub, 1885) Chitwood, 1949 and M. incognita (Kofoid & White, 1919) Chitwood, 1949. Rotation with cowpea and soybean resulted in significant increases in nematode numbers. Maize following cowpea and soybean showed higher nematode populations than both monocultured maize or maize-groundnut rotation sequences. No significant nematode population increases were found in rotations with groundnut. This can be ascribed to the absence of M. arenaria (Neal, 1889; Chitwood, 1949) usually associated with groundnut. M. javanica and M. incognita are generally not associated with groundnut. However, cowpea and soybean, depending on resistance level of individual crop cultivar, are hosts to both species, explaining the significant population increases following rotations with these two crops. Maize yield loss following these rotations increased with an increase in infestation levels of maize roots. Crop rotation in root-knot nematode infested fields with susceptible crops like cowpea, or soybean are not recommended for both commercial and small-scale farming. Key Words: Crop rotation, maize yield loss, Meloidogyne spp., South Africa RÉSUMÉ L'objet de cette étude était d'examiner, sur une période d'essai de quatre saisons, l'effet de la rotation de trois cultures, notamment des graines de soja (Glycine max L. Merr), du dolique (Vigna unguiculata L. Walp) et de l'arachide (Arachis hypogea L.), avec la culture du maïs (Zea mays L.), sur les niveaux d'infestation de nématodes, spécifiquement des anguillules. Les rotations effectuées avec le dolique et les graines de soja ont causé une augmentation considérable de la quantité de nématodes. Quand la culture du maïs a suivi celle du dolique et des graines de soja, les nématodes ont été plus nombreux que dans le cas d'une monoculture de maïs, ou dans les cas d'une succession de maïs et d'arachide. Aucune augmentation importante de la population de nématodes n'a été enregistrée avec les rotations d'arachide. Ce phénomène peut àtre attribué à l'absence de Meloidogyne arenaria (Neal, 1889) Chitwood, 1949; normalement associé à l'arachide. Les espèces dominantes de nématodes d'anguillules présentes dans cet essai, étaient M.javanica (Treub, 1885) Chitwood, 1949 et M. incognita (Kofoid & White) Chitwood, 1949 qui ne sont normalement pas associées à l'arachide. Cependant, le dolique et les graines de soja, compte tenu du niveau de résistance des cultivars individuels de soja, sont des hôtes pour toutes les deux espèces, ce qui explique l'augmentation considérable dans la population de nématodes après la rotation de ces deux cultures. Dans des champs infestés de nématodes, la rotation de cultures susceptibles à ce genre d'infestation, comme le dolique, dans le but de fournir du fourrage vert comme paccage d'hiver aux bovins, ou une culture commerciale avec les graines de soja, ne sont donc pas recommandées. Ceci vaut pour les exploitations commerciales et pour les exploitations à petite échelle. Mots Clés: Réduction de rendement du maïs, Meloidogyne spp., Afrique du Sud INTRODUCTION The presence of nematodes in almost all soil types and cropping systems necessitates consideration in integrated pest management programmes (Dowler and Van Gundy, 1984) particularly in South Africa. Crops often grown by farmers in rotation with maize are soybean (Glycine max L. Merr), cowpea (Vigna unguiculata L. Walp) and groundnut (Arachis hypogaea L.). Increasing input costs and overproduction during wet seasons also lead to uneconomical monoculture production of maize in South Africa. Meloidogyne javanica and M. incognita are the most common root-knot nematode species in South Africa (Kleynhans, 1991) including the western maize production region (Riekert, 1996b). These Meloidogyne spp. are known as parasites of these leguminous crops ( Keetch and Buckley, 1984; De Waele and Jordaan, 1988; Riggs and Niblack, 1993). Keetch (1989) estimated the annual maize yield loss to nematode infestation at 12%, implying a yield loss of 884,520 t. Yield loss to individual species was, however, not calculated. Population build-up of root-knot nematodes and the duration of their life cycle are influenced by soil temperature and moisture (Taylor and Sasser, 1978). At a soil temperature of 26ºC, which is similar to soil temperatures during the maize planting season, the life cycle of M. javanica lasts 21 days (Milne and Du Plessis, 1964). M. incognita occurs also in warmer climates (Taylor and Sasser, 1978). Prolonged drought stress may inhibit development of root-knot nematodes. However, low soil moisture levels of 3.7% still results in survival and root infestation of host crops (Taylor and Sassar, 1978). The dry winter period in production regions will, therefore, provide insufficient nematode control in South Africa. Soil type is important with root-knot nematode infestations. Sandy soils, on which the majority of dryland crops are grown, are prone to nematode infestation and population increases (Van Gundy, 1985). Favourable soil temperature and moisture, as well as soil type and the presence of Meloidogyne spp. may, therefore, result in yield reduction in susceptible crops. This study was aimed at determining the influence of cowpea, groundnut and soybean rotations with maize as main crop on root-knot nematode population levels and the effect of infestation levels on maize following these crops in a M. javanica and M. incognita infested field. MATERIALS AND METHODS A field trail was conducted at Skandinawiâdrif, in the northern Free State Province of South Africa, from the 1992/93 to the 1995/96 planting seasons. Plots were prepared using a mouldboard plough since the soil had a clay content <10%. Maize (Cultivar: PAN6043) was planted both in rotation and mono-culture for four consecutive seasons. Rotations included both one (in the third season) and two rotations (in the first and fourth season) within four seasons with either cowpea, (Cultivar: Glenda), groundnut (Cultivar: Sellie) and soybean (Cultivar: Knap). The rotations are depicted in Table 1. Plot size was 6.3 m x 10 m. A randomised complete block design was used, with all cropping sequences replicated three times. Initial Meloidogyne population levels in the trial site, previously under monoculture maize cultivation, ranged between 0 and 50 eggs and larvae with most counts less than 20 in 50 g roots. TABLE 1. Rotation schedule of individual plots within the randomised complete block trial layout
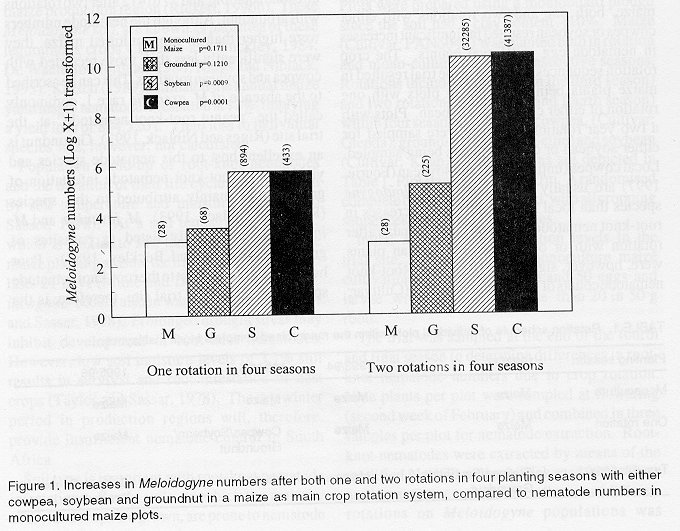
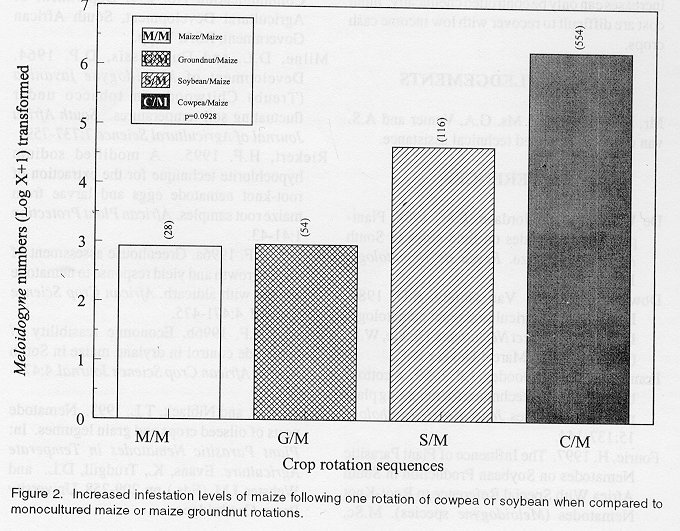
The trial was sampled at the end of the fourth and final season to determine differences in root-knot nematode numbers due to crop rotation. Nine plants per plot were sampled at flowering (second week of February) and combined in three samples per plot for nematode extraction. Root-knot nematodes were extracted by means of the modified NaOCI-method (Riekert, 1995). Eggs and larvae were counted. The influence of different rotations on Meloidogyne populations was determined by comparing nematode counts of individual crops to that of monocultured maize. The effect of crop rotations on infestation levels in maize was done by comparing nematode numbers in maize plots following either cowpea, soybean or groundnut with counts from monocultured maize. Nematode numbers were (log X + 1) transformed and subjected to analyses of variance. Species identification was done by removing 30 adult females from separately collected root samples from all plots in the trial. Roots were stained, using the lactophenol-acid fuchsin method (Franklin and Goody, 1959) to facilitate dissecting. Perineal pattern identification was done after preparation of the patterns according to the procedure of Hartman and Sasser (1985). RESULTS AND DISCUSSION Nematode numbers. M. javanica and M. incognita were the only root-knot nematodes present at the trial site and occurred in a ratio of 7:3, respectively. Compared with monoculture maize, both one and two rotations with either cowpea or soybean resulted in significant increases in nematode egg and larval counts. The crop rotation sequences followed in the trial resulted in maize plants being sampled in plots with one rotation of either cowpea or soybean. Plots with a two year rotation sequence were sampled for either soybean or cowpea plants being sampled. Local cowpea (unpubl. data) and soybean (Fourie, 1997) are usually better hosts to Meloidogyne species than local maize cultivars. Increases in root-knot nematode numbers, both on maize after rotation with or on cowpea or soybean plants were, however, significant (P<0.05). Root-knot nematode counts of monocultured maize (Cultivar: PAN 6043) compared to one rotation and two rotations of cowpea (Cultivar: Glenda) are provided in Figure 1. Both these rotations resulted in significant increases in the nematode population. One rotation with cowpea resulted in counts ranging from 83 to 861 eggs and larvae in 50 g root samples (maize plants sampled). Counts after two rotations (cowpea plants sampled) ranged from 15,690 to 73,600 eggs and larvae compared to an average of 28 eggs and larvae in monocultured maize. Rotations with soybean (Cultivar: Knap) also resulted in significant (P<0.0009) increases in nematode numbers compared with monocultured maize (Fig. 1). One rotation resulted in counts from 48 to 3,917 (maize plants sampled), while two rotations resulted in counts ranging from 20,746 to 51,616 (soybean plants sampled). Groundnut rotations (Cultivar: Sellie) did not result in significant (P=0.121) increases in root-knot nematode numbers compared to monocultured maize (Fig. 1). Counts in 50 g root samples ranged from 0 to 112 eggs and larvae after one rotation, and 77 to 412 after two rotations with groundnut. Although the nematode numbers were higher than in monocultured maize, they were significantly less than those recorded with cowpea and soybean rotations. This can be ascribed to the absence of M. arenaria race 1, commonly called the "peanut root-knot nematode" at the trial site (Riggs and Niblack, 1993). Groundnut is an excellent host to this nematode species and yield loss to root-knot nematode infestation of groundnut is mainly attributed to this species (Riggs and Niblack, 1993). M. javanica and M. incognita are locally listed as parasites of groundnut (Keetch and Buckley, 1984). Poor host status of groundnut to the root-knot nematode species present at the trial site, therefore, is the causal factor in the observed tendency when compared with cowpea and soybean. This also explains the impact of these rotation crops on the infestation of maize. Due to a trial duration of four seasons, only one rotation effect on maize as a follow-up crop of each of the rotation crops was possible. An increase (P=0.0928) in infestation levels of maize (average 116 and 554 in 50 g of roots, respectively) was obtained after one rotation of soybean and cowpea (Fig. 2) compared with monoculture maize (28 in 50 g roots). Despite being nonsignificant (P=0.0928), the fourfold increase in root-knot nematode infestation levels of maize obtained after one rotation with soybean and the twentyfold increase after one rotation with cowpea are regarded as important. This is the direct result of the significant increases in nematode numbers on these crops as presented in Figure 1. Groundnut rotations resulted in nonsignificant increases of infestation levels in maize. This can be explained by the poor host status of groundnut to the Meloidogyne population present compared to the host status of cowpea and soybean. Monoculture maize (Cultivar: PAN6043) resulted in the lowest egg and larvae counts 50 g roots-1 with a range of 0 and 77 per sample and an average of 28. Four year monoculturing of maize resulted in nonsignificant root-knot nematode increases at this trial site compared with nematode numbers in soybean and cowpea root samples (Fig. 1). However, this does not imply that monoculture maize does not result in root-knot nematode build-up. In a field trial at Viljoenskroon with the same soil type, infestation levels in monocultured maize increased from less than 10 eggs and larvae 50 g roots-1 to as high as 11,000 eggs and larvae (Unpublished data). M. javanica and M. incognita ratio were present in a 9:1. This was inspite of more favourable environmental conditions especially rainfall. FIGURE 1: Increases in Meloidogyne numbers after both one and two rotations in four planting seasons with either cowpea, soybean nd groundnut in mize as main crop rotation system, compared to nematode numbers in monocultured maize plots. Yield data. Yield data of the fourth season are included in order to emphasise the importance of prevention of population build-up (Fig. 2). Comparison of yield data over seasons was not possible due to variations in annual rainfall combined with differences in drought tolerance of the various crops. The increased root-knot nematode infestation levels of maize following the rotation crops resulted in maize yield loss. The average yield of the monoculture plots was 4,782 kg ha-1. An increase in nematode egg and larvae numbers from 28 to 54 in 50 g roots after rotations with groundnut resulted in a maize yield of 2,690 kg ha-1. Maize yield further declined to 2176 kg ha-1 following soybean and an infestation level of 116 eggs and larvae in 50g maize roots. Infestation levels of 554 eggs and larvae in 50 g maize roots following cowpea provided a yield of 1,898 kg ha-1. Although some yield loss was possibly due to increased water use by the crops preceding maize, it is important to note a decrease in yield with each increase in infestation, compared to maize in monoculture. FIGURE 2: Increased infestation levels of maize following one rotation of cowpea or soybean when compared to monocultured maize or maize groundnut rotations. SUMMARY Cultivation of the cowpea cultivar Glenda and soybean cultivar Knap resulted in increased M. javanica and M. incognita populations and increased infestation levels of the maize cultivar PAN6034 resulting in yield loss. Host suitability combined with root-knot nematode species composition at the trial site were causal factors for significant increases in infestation levels of maize and subsequent yield loss. Since chemical control of nematodes in dryland cultivated maize is seldomly economically viable (Riekert, 1996b), population build-up of plant parasitic nematodes should be prevented by other means. M. javanica and M. incognita were species in the nematode complex, resulting in a 50g plant-1 yield loss to maize in a greenhouse trial (Riekert, 1996a). These levels of population increases thus have negative economic and yield effect on maize production. Screening of local soybean cultivars are presently being conducted at the Grain Crops Institute. Differences in resistance between cultivars were recorded, emphasising the importance of cultivar selection in crop rotation (Fourie, 1997). Local cowpea cultivars and lines are also presently being screened but no differences in host suitability is yet obtained. The nematode species composition seem to influence the effect of individual crops in a crop rotation system, as emphasised in this study. The absence of M. arenaria resulted in lower infestations and population increases in plots rotated with groundnut, but the presence of M. javanica and M. incognita had an adverse effect on plots with cowpea and soybean rotations. To plan successful crop rotation systems in an integrated pest management system, especially with low income crops under dryland cultivation, nematode populations present in the soil and host status of crops or cultivars should be taken in consideration. High nematode population increases can only be controlled chemically. Input cost are difficult to recover with low income cash crops. ACKNOWLEDGEMENTS Mr. M.J. Schoeman, Ms. G.A. Venter and A.S. van der Walt provided technical assistance. REFERENCES
Copyright 1998, African Crop Science Society The following images related to this document are available:Photo images[cs98040b.jpg] [cs98040a.jpg] |
| |||||||||

{kind=link}
{kind=link}