 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, SHORT COMMUNICATION Classification Of Sorghum Germplasm Accessions Using Multivariate Methods I.E. EZEAKU, S.C. GUPTA and V.R. PRABHAKAR ICRISAT, P M B 3491, Kano, Nigeria (Received 21 April, 1998; accepted 26 October, 1998) Code Number: CS99010 ABSTRACT Two multivariate methods were used to classify 352 accessions of sorghum (Sorghum bicolor (L.) Moench) originating from Nigeria and Chad. The study revealed the existence of a considerable range of variability within the cultivated sorghums for 13 traits studied. Days to flowering represented 64.8% of the total variation observed within the 352 accessions. Based on flowering, three groups were formed consisting of early, medium and late flowering accessions. Eight clusters were formed in each flowering group following Mahalanobis D2 analysis. The accession numbers in each cluster are listed for possible seed requests from sorghum breeders. Principal components analysis indicated that plant height, panicle length and floaters (%) contributed most towards genetic divergence among the accessions studied. Clustering pattern revealed that geographic diversity, though important, is not the only factor responsible for determining genetic divergence. The implications of this study to sorghum breeding are summarised. Key Words: Genetic diversity, Mahalanobis D2, ICRISAT, Nigeria, Sorghum bicolor RÉSUMÉ Deux méthodes multivarietales étaient utilisées pour classer 352 accessions du sorgho (Sorghum bicolor (L.) Moench) en provenance du Nigeria et du Tchad. Cette étude a révelé l=existence d=une variabilité considérable parmi les sorgho cultivés pour les 13 traits étudiés. Après quelques temps à la floraison, ils ont présenté 64,8% de la variation totale observée parmi les 352 accessions. Basé sur la floraison, trois groupes ont été formés pour observer les accessions précoces, moyennes et tardives. Huit clusters ont été formés au sein de chaque groupe de floraison suivant l=analyse de Mahalanobis D2. Les nombres d=accession dans chaque composante sont énumerés pour satisfaire aux demandes de semences des selectionneurs de sorgho. L=analyse des composantes principales ont indiqué que la taille des plants, la longueur des panicules, et le pourcentage des grains flottants ont contribué le plus à la divergence génétique parmi les accessions étudiées. Le régime des composantes a révelé que la diversité géographique, bien qu= importante, n=est pas le seul facteur responsable pour déterminer la divergence génétique. Les implications de cette étude sur la sélection du sorgho sont résumées. Mots Clés: Composantes principales, diversité génétique, Mahalanobis D2, multivariétale, ICRISAT, Nigeria, Sorghum bicolor INTRODUCTION Sorghum varieties were traditionally classified by Harlan and de Wet (1972) into five races (bicolor, caudatum, durra, guinea, and kafir) on the basis of morphological traits. House (1985) noted that cultivated sorghums are highly variable. Doggett (1965) speculated that ethnic isolation resulted in morphological differences between races. Similar views have been expressed by Simmonds (1986), although he added that domestication as well as eco-geographic diversity accounted for changes in several features of plants resulting in diverse populations. In a recent survey, Appa Rao et al. (1994) reported that Nigerian Kauras and Fara-fara possess a wide range of variation, especially in panicle and grain traits. In support of this observation, it was found that farmers in northern Nigeria call sorghums Badari (yellow grain), Goronya (gooseneck), Rosuba (chalky white grain) or Mori (normal white grain). Increases in sorghum yield derived from genetic manipulations depend on the existence of appropriate genetic variability (House, 1985). Sabharwal et al. (1995) emphasised that sorghum parents with more diversity among them are expected to exhibit a higher amount of heterotic expression and a broad spectrum of variability in segregating generations. Deu et al. (1994) suggested that multivariate techniques may be helpful in quantifying the degree of relatedness among landraces. Despite some shortcomings of multivariate techniques, they have allowed efficient varietal classification for many crops. These techniques have been widely used in the classification of inbred lines and accessions within sorghum (Shanmuga-sundaram and Subramanian, 1990; Vierling et al., 1994; Sabharwal et al., 1995), Triticum spp. (Vierling and Nguyen, 1992; Monte et al., 1993), maize (Livini et al., 1992), and soyabeans (Ezeaku and Awopetu, 1992). These techniques group operational taxonomic units (OTUs) on the basis of overall similarity or dissimilarity using a large number of characters (Sokal and Sneath, 1963). It was observed by Chheda and Fatokun (1982) that these methods enable the division of a vast number of strains into clearly discernible categories which cannot be handled by other classification methods. Computer programmes of these techniques permit genotypes to be classified and incorporated either into appropriate existing groups or to be placed into newly created groups by re-running the computer programme on the accumulated data. Rhodes et al. (1969), in support of these techniques, stated that when the number of entries in a gene bank must be curtailed due to financial and spatial constraints, the choice of those to be eliminated will be aided by such a classification system. This study is aimed at elucidating the genetic affinity within the germplasm collected during 1995, in order to provide a basis for selection of parents for hybridisation. MATERIALS AND METHODS This study involved 352 sorghum [Sorghum bicolor (L.) Moench] accessions including 338 from northern Nigeria, and 14 from Chad. These accessions were collected during 1995 by the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) and the Lake Chad Research Institute (LCRI). In Nigeria, accessions were collected from 11 states. These 352 accessions were sown at Bagauda, Nigeria on June 27, 1996. Bagauda is located at 11º 53' N latitude and 8º 14' E longitude at a mean altitude of 440 m above sea level. Soil type is a plinthic luvisol with an average depth of 90 cm. Seeds of each accession were sown in two rows of 5 m. The plant to plant spacing between rows was 75 cm and within-row 25 cm. The experiment was sown unreplicated (due to a large number of entries), in a uniform piece of land. Fertilizer (NPK 15:15:15) was applied basally at a rate of 300 kg ha-1 by uniform hand-broadcasting and then incorporated into the soil. Three weeks after sowing, calcium ammonium nitrate (CAN) was applied at 100 kg ha-1. CAN was placed about 5 cm from the plant base at a depth of about 3 cm in furrows made with a furrow opener. The fertilizer was then covered to avoid losses through volatilisation. Inter-culture and hand-weeding were done when necessary. The data on 12 plant traits (Table 1) were recorded using the International Board for Plant Genetic Resources (IBPGR) and ICRISAT Descriptor list for sorghum (IBPGR/ICRISAT, 1993). To estimate floaters (%), a solution was prepared by dissolving 667 g of sodium nitrate crystals in one litre of distilled water. The percent of seeds floating on the solution was expressed as floaters (%).The first step employed in this analysis was to limit the 352 OTUs to a manageable size by categorising them into three distinct flowering groups of early (60 to 87 days), medium (90 to 116 days) and late (117 to 162 days) using Mahalanobis (1936) D2 statistics. Mahalanobis D2 statistics and principal components analysis were carried out using Genstat 5 statistical package (Genstat 5 Committee, 1989). Accessions were classified into different clusters by using Tocher=s method (Rao, 1952). RESULTS By using Tocher>s method as described by Rao (1952), days to flowering (half bloom) represented 64.8% of the total variation observed within the 352 sorghum accessions studied, while the remaining 12 traits accounted for 35.2%. Relatively high variation for this trait indicated a greater contribution to phenetic diversity. These 352 OTUs were divided into three distinct flowering groups of early, medium and late by using D2 statistics. The accessions in each flowering group were further subdivided into eight clusters by using the same statistics. The characteristics of these eight clusters for each flowering group are presented in Table 1. The number of accessions in clusters varied from 1 (cluster V) to 40 (cluster VI) in the late flowering group. In the early flowering group, cluster VI possessed the largest number of accessions (18), and cluster IV had the least (7). In the medium flowering group, cluster VIII possessed the highest number of accessions (20), and cluster V had only 5 accessions (Table 2). Most of the accessions in all flowering groups had non-tan plant colour except a few accessions in the early flowering group (such as entry 17 - SG 909, cluster VIII, Figure 1). SG 909 could be used to transfer tan plant trait into selected local landraces. All accessions of cluster V in all flowering groups possess plump grain features. On average, the grains of the medium flowering accessions were heavier than those of early and late flowering accessions (Table 1). This group could provide source material for development of bold-seeded hybrids and varieties. Most medium and late flowering accessions are of semi-loose head shape. TABLE 1. Description [mean (standard deviation)] of the eight clusters produced by Mahalanobis D2 analysis of Early (E), Medium (M), and Late (L) flowering sorghum accessions
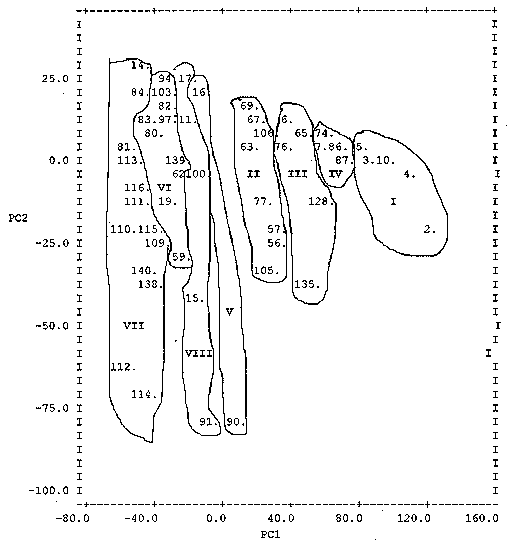
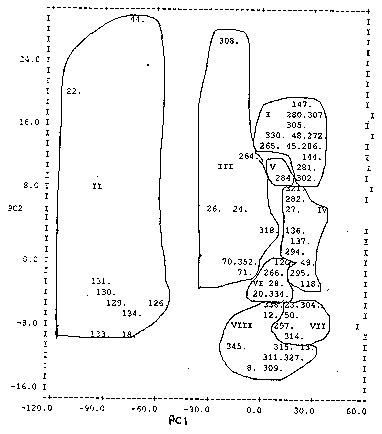
Seventy eight percent of the early flowering accessions were materials collected from Katsina and Sokoto states in Nigeria. These states are characterised by low rainfall of short duration. Similarly, 81% of the accessions of the late flowering group were collected from the Federal Capital Territory (Abuja), Niger, Bauchi and Plateau states, also in Nigeria. These states have higher and longer rainfall duration than Katsina and Sokoto states. This pattern of distribution of accessions may be due to adaptation and natural or human selection. On the other hand, the accessions of medium flowering type grouped independently across all the states where collections were made, reflects the broad spectrum of adaptation potentials of these accessions. All the introductions from Chad clustered independently within and across the three flowering groups, indicating a high level of variability in these introductions. The 352 accessions divided into three flowering groups and eight clusters in each group are listed in Table 2. These accessions are coded using SG numbers and are kept in temporary storage at ICRISAT, Kano, Nigeria. Clusters within the medium and late flowering groups contain the most related accessions, while clusters within the early flowering group contain the least related accessions. Clustering pattern of accessions in Table 2 (Mahalanobis, D2) follows similar pattern with the principal components (Figures 1, 2 and 3). Figure 1: Ordination of early flowering accessions showing general scatter and grouping along principal components 1 and 2. Figure 2: Ordination of medium flowering accessions showing general scatter and grouping along principal components 1 and 2. Figure 3: Ordination of late flowering accessions showing general scatter and grouping along principal components 1 and 2. TABLE 2. Classification of 352 sorghum accessions into clusters/flowering groups using SG numbers as identifiers
The intra- and inter- cluster D values for eight clusters of each flowering group are presented in Table 3. Maximum inter-cluster divergence occur between clusters I and VII, clusters I and II and clusters I and V for early, medium and late flowering groups, respectively. The inter-cluster divergence was least between clusters III and IV of early flowering group, VI and VIII of medium flowering group and VI and VII of late flowering group. Based on observations from intra- cluster range, accessions in the medium and late flowering groups appear to be more related than those in the early flowering group. The crosses made between accessions of clusters I and VII (early), clusters I and II (medium) and clusters I and V (late) are expected to generate greater heterotic effect than the crosses made between accessions of clusters III and IV (early), VI and VIII (medium) and VI and VII (late). Similar results are obtained by principal components (PC) analysis (Figures 1, 2 and 3). The D value has helped to identify the diverse accessions from the available germplasm, which will be useful in future breeding work. Principal components analysis was conducted to represent the diversity in a lower dimensional space for better understanding of the major sources of diversity and the inter-relationships among the accessions . The set of 12 variables measured, excluding days to flowering, were reduced to two principal components which are linear combinations of the original variables. Principal components were computed for each flowering group separately after standardisation of the original variables. The first two principal components explained 77.78 and 19.23% in the early flowering group, 84.22 and 8.32% in the medium, and 52.27 and 29.34% in the late flowering group. The elements of the principal component vectors differed both in sign and magnitude among the flowering groups. This indicates that the variables that discriminate the accessions differ among the flowering groups. TABLE 3. Inter-cluster and intra-cluster (in bold) D values among eight clusters of each flowering class
The ordination of the 86 early, 107 medium and 159 late flowering accessions along the axis of PC 1 and PC 2 are presented in Figures 1, 2 and 3, respectively. These two components were plotted with entry numbers as identification points. The graphs clearly depict the clusters although the boundaries separating the clusters are not very clear in some cases. The loading of the latent vectors for the clusters (Table 4) indicates the importance of the variables in making the components. The loadings for the early flowering group reveal that the floaters (%), panicle length and plant height generally dominate the principal components. These traits explain most of the genetic diversity of the early flowering group. This implies that the accessions from different clusters in the early flowering group differ mostly with respect to these traits. For instance, in Figure 1, accession 14 (SG 906) has a high PC2 value of 100% floaters, while accessions 90 (SG 982) and 91 (SG 983) have a low PC2 values with 7% and 4% floaters, respectively. Similarly, in the late flowering group, plant height and floaters (%) dominate the principal components (Table 4). A high value on the PC1 axis indicates that seeds are heavy and a high value on PC2 indicates that the accession is tall. TABLE 4. Latent vectors (loadings) for 12 traits in grain sorghum
PC = Principal component In the medium flowering group, plant height, panicle length and floaters (%) contributed most to the genetic diversity. A high value on PC1 axis indicates that the accession is tall and a high value on PC2 axis indicates long panicle and low floaters (%). The accession 308 (cluster III) and 309 (cluster VIII) have similar PC1 but 308 (SG 1200) has a high PC2 value and 309 (SG 1201) has a low PC2 value (Figure 2). This shows that both accessions are tall but 308 has a much longer panicle than 309. This observation is corroborated by data in Table 1. For example, accessions in cluster III and cluster VIII in Figure 2 are all tall with a mean plant height of 347 and 367 cm, respectively. Conversely, accessions in cluster III possess longer panicles (mean panicle length, 36 cm) than those in cluster VIII (mean panicle length, 23 cm). Accessions in Figures 2 and 3 appear more homogeneous than accessions in Figure 1. Accession 207 appears to be the only non-conformist in Figure 3. This accession falls into cluster V with only one accession (Table 1 and Figure 3). It is characterised by exceptionally long panicle, and could be used as a source material in a breeding programme. DISCUSSION The two multivariate procedures of non- hierarchical clustering (Mahalanobis, D2) and principal components used in this study are complementary since information provided by both techniques are, to a large extent, similar. El Gazar et al. (1986) postulated that the position of OTUs in different taxonomic procedures, when using the same set of data, may be due to the taxonomic characteristics of the respective methods. Our results indicated that eco-geographical factor, though important, appears not to be the only factor in determining germplasm accession distribution. This is contrary to the findings of Sabharwal et al. (1995) who concluded that there was no association between clustering pattern and eco-geographical distribution of accessions. It is also at variance with the findings of Joshi and Dhawan (1966) and Rao and Paroda (1982) who advocated that eco-geographical distribution has control over genetic diversity. The studies on genetic divergence in sorghum conducted by Shanmugasundaram and Subramanian (1990) revealed that factors other than eco-geographical region contributed significantly to genetic divergence which is confirmed by the present findings. The observed relationship between centre of collection and accession grouping suggests that it is necessary to collect and conserve germplasm representing a broad geographical range to capture as much diversity as possible. This would enhance the development of sorghum ideotypes. Bekele and Bekele (1996) noted that this link may have implications in the identification of accessions for a core collection and, thus warrants further investigation as a preamble to the assessment of allelic diversity to establish such a core collection. Days to flowering has contributed most to the genetic diversity in the accessions studied. Within flowering groups, the major contribution to the genetic diversity was due to plant height, panicle length and floaters (%) (Table 4). Shanmuga-sundaram and Subramanian (1990) reported that grain yield, straw yield, grain number, peduncle thickness, grain weight, leaf number, panicle length and days to half bloom contributed the most to genotypic differences. Murthy et al. (1967) found that spikelet number, distance between whorls, panicle length and days to flower were major contributors to genetic divergence. Our study further reveals variability in grain shape, size, colour and in inflorescence characteristics. Late flowering accessions were found to exhibit semi-loose head shape. This is consistent with the findings of Appa Rao et al. (1994). All the accessions sampled expressed high incidence of foliar diseases, exhibiting narrow genetic base for these traits. Simmond (1986) stated that leaf diseases are always present but rarely epidemic in landrace germplasm. Higher genetic variability is likely to be created when the crosses are made between selected parents from different clusters than within cluster (Gardner, 1972; Fatokun, 1985; Ezeaku and Awopetu, 1992). The useful genes for several traits such as earliness, short stature, long and thick panicles, bold and dense seeds, and tan plant colour exist within the germplasm accessions studied. These genes can be incorporated to improve sorghums of each flowering group. Most of the sorghums grown in Nigeria are of medium to late flowering types. This is evident from germplasm accessions collected in 1995 (Table 1). These sorghums are cultivated by small-holder farmers as a component of mixed cropping, and suffer from terminal drought. The flowering period of these sorghums can be reduced by crossing with short duration accessions. Short stature types can be found in cluster I of the early flowering group and could be good potential parents for hybridisation since dwarfing genes could be unlocked from them. Early flowering accessions are mostly small-seeded. The seed size in early flowering accessions can be improved by introducing genes from accessions of the medium flowering group (clusters II and VIII). Most of the accessions had non-tan plant colour, indicating that non-tan types are most adapted to Nigerian environments. Similarly, semi-loose head shape was widely observed in medium and late flowering accessions. Such traits are perhaps a result of selection and adaptation, and therefore, should be preserved in future breeding programs. ACKNOWLEDGEMENTS The authors are grateful to Dr. R. Tabo for collecting germplasm from Chad, and to Messrs Yusuf Ali (LCRI) and S. E. Aladele for collecting germplasm from Nigeria. We are also grateful to Drs. Subhash Chandra and O. Ajayi for reviewing this manuscript. REFERENCES
Copyright 1999, African Crop Science Society The following images related to this document are available:Line drawing images[cs99010b.gif] [cs99010c.gif] [cs99010a.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}